

As global forests wrestle with the escalating threats of climate change, the study of forest regeneration, particularly through the lens of tree reproduction, becomes increasingly important. For forest trees, the three fundamental life processes are growth, mortality, and reproduction. Yet, we know much less about the last one than we do about the first two. Tree reproduction is more than a natural process — it's the lifeline that ensures the continued survival of our forests. In the era of significant global environmental changes, there is a pressing need to understand how tree reproduction can potentially counterbalance the rising rates of tree mortality. Furthermore, tree reproduction plays a pivotal role in shaping the forest food web. The variations in seed production directly influence the availability of food for seed consumers, which can trigger cascading effects on population dynamics and interactions among various species in the ecosystem.
Tree reproduction is a complex process that becomes even more intriguing due to the phenomenon known as "masting”. This term describes the significant fluctuation in seed production, not just between individuals of the same species but also from one year to another within a single tree — a variation that can span orders of magnitude. Such variation, which presents low signal-to-noise data, demands extensive sample sizes to estimate seed supply. A global database known as the "Masting Inference and Forecast" (MASTIF) has been compiled through an international collaboration led by Professor James S. Clark from Duke University, involving researchers from over 70 institutions worldwide. This comprehensive database consists of more than 12 million observations, serving as a foundational tool to quantify global seed supply and its response to global change.
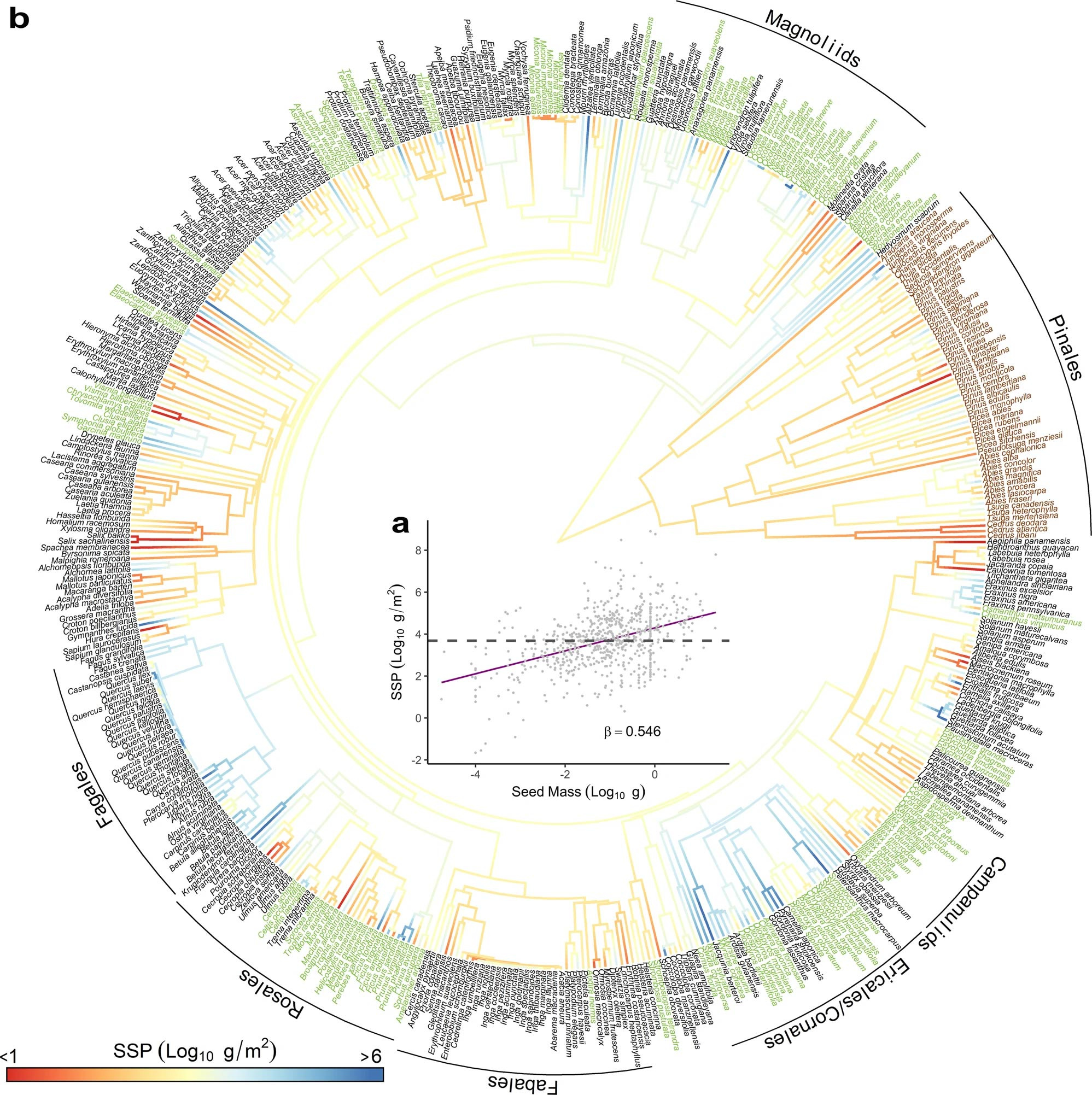
The MASTIF database allows us to test the long-debated hypotheses concerning the trade-off between seed size and numbers. For years, ecologists have observed an intriguing strategic decision trees make: they can choose to generate a limited number of large seeds, each packed with enough resources to enhance seedling survival, or opt to produce an abundance of small seeds. These smaller seeds may compensate for their lower survival rates by increasing the chances of reaching suitable sites for growth. While the variations in seed size across different species have been widely observed and extensively studied, the same cannot be said for seed numbers. This is an essential element in understanding the evolution of tree species and how they respond to global forest dieback. To fill this knowledge gap, we introduced a new concept: species seed production (SSP). SSP is calculated as the product of seed number and seed size. We used this measure to capture the variations of SSP across more than 700 species globally. If the trade-off theory holds true, SSP would show limited variation across species. However, our analysis uncovered a new trend: big-seed species tend to generate more seeds, leading to ten order-of-magnitude differences in SSP (Fig. 1). Furthermore, we found gymnosperms, also known as conifers, have lower SSP than angiosperms, or the flowering trees (Fig. 1). This could be attributed to the substantial energy conifers invest in creating protective cones for their seeds. These findings not only hold practical implications for forest management but may also shed light on Darwin's famous "abominable mystery" — the rapid diversification of flowering plants during the Cretaceous period. The ability to produce a large number of seeds may have been the critical factor that enabled these flowering plants to flourish and evolve under the harsh conditions of the Cretaceous period. This insight seems to hold relevance today, just as it did millions of years ago. These results were published in Nature Communications in the year 2022 https://www.nature.com/articles/s41467-022-30037-9/.
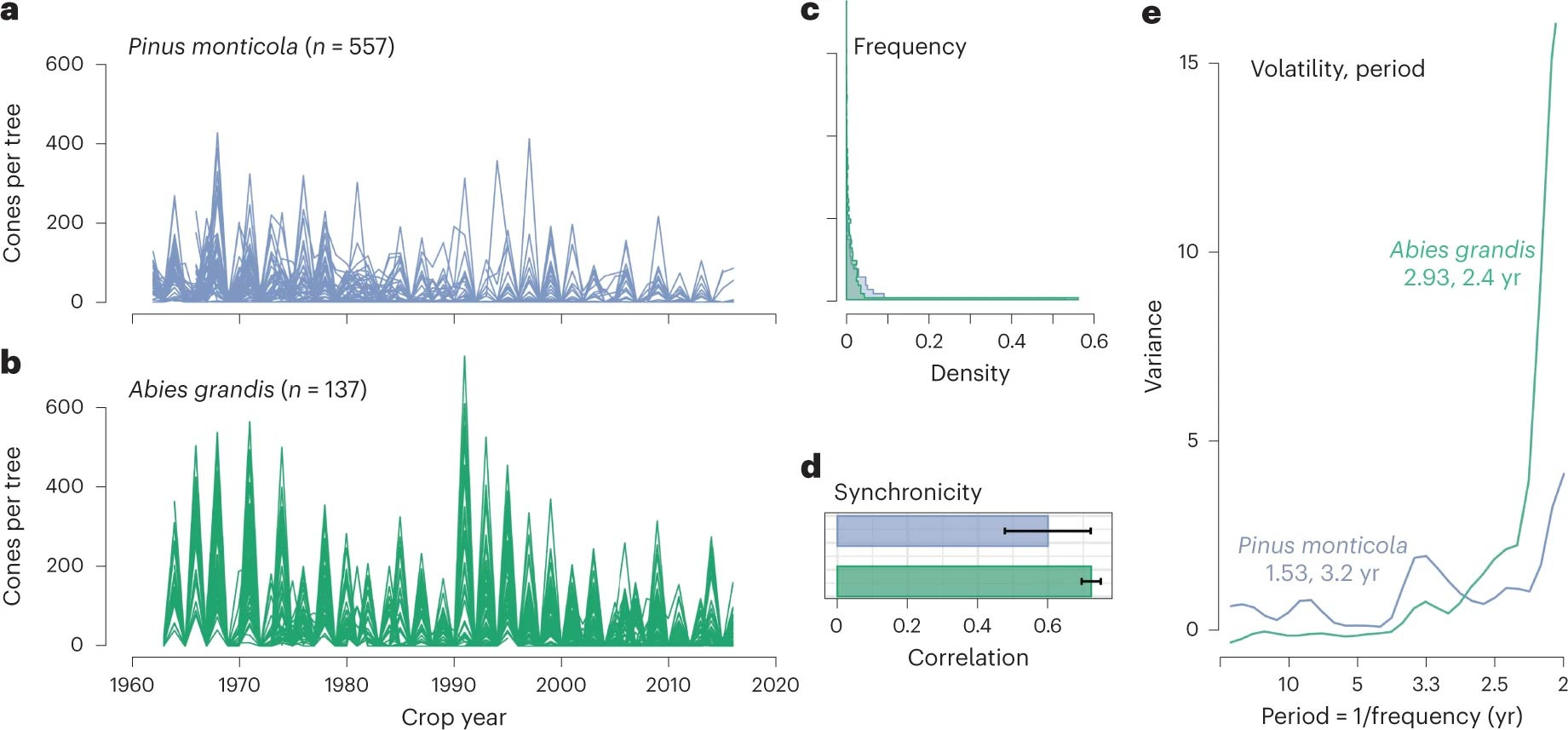
After revealing the patterns and implications of mean seed supply through species seed production (SSP), we turned our attention to understanding the year-to-year variations in seed production, or masting. The phenomenon, characterized by significant fluctuations in interannual seed production, long intervals between high seed years, and synchronized production of abundance seeds, can potentially overwhelm seed predators. This reproduction strategy limits the consumer’s ability to mitigate interannual variations in one host tree by foraging from others. But it raises the question: wouldn’t the unreliable seed production that thwarts a tree’s foes also adversely affect their mutualist disperser? Furthermore, could the highly synchronized seed production intensify competition among offspring? To answer these questions, we introduced three elements that offer new ways to jointly measure masting (Fig. 2): 1) volatility, illustrating the dynamic nature of seed production from year to year — not a steady flow, but a series of peaks and troughs that keeps predators guessing; 2) periodicity, indicating the time lapse between years of high seed supply that leaves predators in a continuous state of uncertainty; 3) synchronicity, representing the phenomenon that trees, even across long geographical distances, seem to be in tune with each other and produce large seed crops simultaneously. We then evaluated the mixed benefit of masting on seed consumers and dispersers within the context of different environmental conditions.
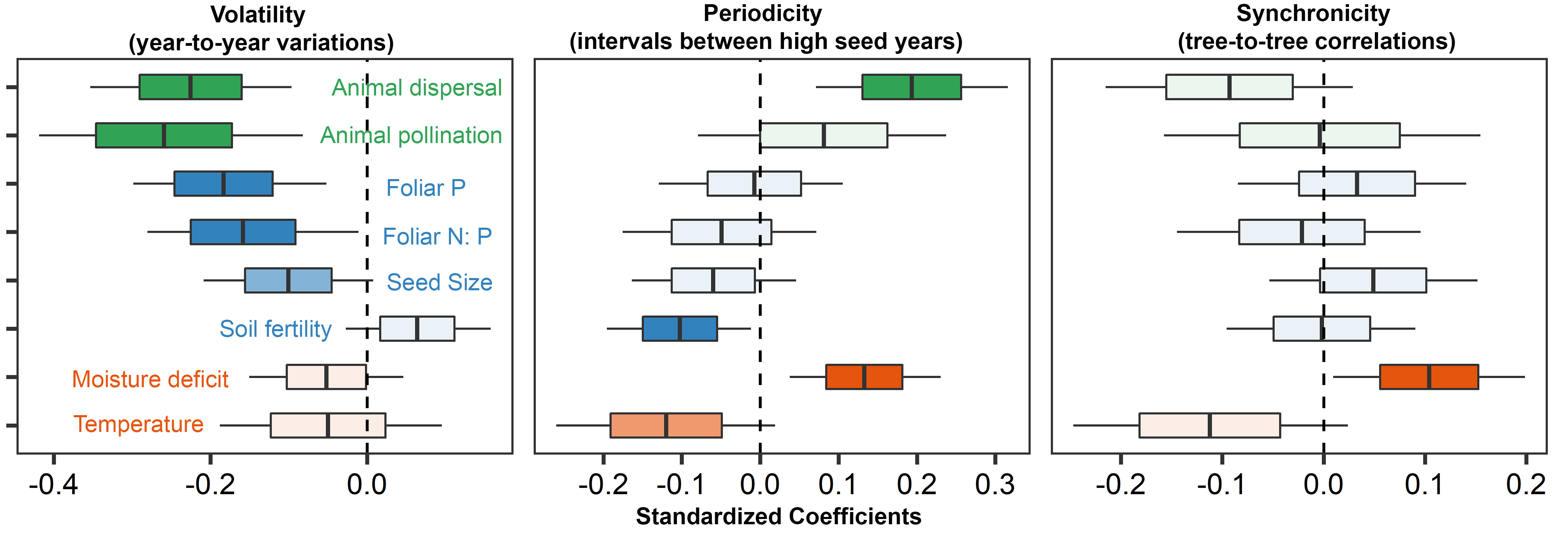
Results show that tree species most reliant on animal dispersers tend to avoid masting (negative coefficient in Fig. 3). Tree species in temperate forests of North America and Eurasia, such as firs and oaks, are well-known for their extensive masting. Pines and spruces also mast, but to a lesser degree. By contrast, trees producing rich, colorful fruits tend to be more “reliable” food sources due to their heavy reliance on animal dispersers. Nutrient availability and climate are additional factors influencing masting (blue and red boxes in Fig. 3). Specifically, tree species with high nutrient demands typically exhibit less volatility, and species observed in nutrient-rich, warm, and wet regions often have shorter intervals between high seed years. Masting is more commonly observed in colder and drier regions because the roles of seed dispersers are less pronounced than in wet tropics. This intriguing interplay underscores the fact that our diverse forests are the product of numerous interacting factors, with trees continually adapting to their unique environmental conditions to thrive.
Acknowledgment: This research would not have been possible without the support of many data contributors in the MASTIF network. The MASTIF project has been funded by the National Science Foundation (NSF) and the National Aeronautics and Space Administration (NASA) to PI James S. Clark.
Other References on the MASTIF project:
Clark, J. S., et al. "Foodwebs based on unreliable foundations: spatiotemporal masting merged with consumer movement, storage, and diet." Ecological Monographs 89.4 (2019): e01381. DOI: https://doi.org/10.1002/ecm.1381
Clark, J. S., et al. "Continent-wide tree fecundity driven by indirect climate effects." Nature Communications 12.1 (2021): 1242. DOI: https://doi.org/10.1038/s41467-020-20836-3
Qiu, T, et al. "Is there tree senescence? The fecundity evidence." Proceedings of the National Academy of Sciences 118.34 (2021): e2106130118. DOI: https://doi.org/10.1073/pnas.2106130118
Qiu, T, et al. "Limits to reproduction and seed size-number trade-offs that shape forest dominance and future recovery." Nature Communications 13.1 (2022): 2381. DOI: https://doi.org/10.1038/s41467-022-30037-9
Journé, V, et al. "Globally, tree fecundity exceeds productivity gradients." Ecology letters 25.6 (2022): 1471-1482. DOI: https://doi.org/10.1111/ele.14012
I am a broadly trained ecologist interested in understanding the response of the terrestrial ecosystem to global change at regional to global scales. My lab takes a data-model synthesis approach that integrates satellite and airborne remote sensing, monitoring networks, and forest inventory with Bayesian hierarchical models to develop nature-based solutions to fight climate change. We are interested in understanding tree reproduction variations and how they relate to climate, nutrient availability, and habitat conditions. We are also interested in understanding the complex interactions between climate warming and habitat that govern biodiversity change in multiple species groups.
Follow the Topic
-
Nature Plants
An online-only, monthly journal publishing the best research on plants — from their evolution, development, metabolism and environmental interactions to their societal significance.




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in