Antarctic haloarchaea – a unique Microbiome
Published in Microbiology

The paper in Microbiome is here: https://rdcu.be/1h8T
Haloarchaea are members of the domain Archaea that thrive in hypersaline environments. In Antarctica, the most well characterized haloarchaea come from a deep (36m), cold (as low as -20°C), marine-derived system called Deep Lake (DeMaere et al PNAS 2013). The lake is home to a low complexity community of microorganisms that is dominated by species of haloarchaea (Halohasta litchfieldiae [tADL], DL31 and Halorubrum lacusprofundi) that don’t tend to dominate in lower latitude systems. By environmental microbiology standards (where the majority of indigenous microbes are recalcitrant to laboratory isolation attempts), Hrr. lacusprofundi is exceptional in being amenable to isolation and cultivation. The ability to obtain new strains of the species have already proven to be of immense value with strain R1S1 providing the discovery of the plasmid (pR1SE) that ‘masquerades’ as a virus and disseminates via membrane vesicles (see Nature Research Microbiology Community); Erdmann et al Nat Microbiol 2017).
The Antarctic haloarchaea from Deep Lake have also been found to be quite ‘promiscuous’, capable of sharing long (up to 35kb), high identity (~100% nucleotide identity) regions (HIRs) of DNA (DeMaere et al PNAS 2013). The ability to do so has interesting implications for the ‘pan-genome’ of haloarchaeal species – i.e. the total pool of genetic material comprised by all members of a species (Tettelin et al PNAS 2005). Given the dominant Deep Lake Antarctic haloarchaeal species differ to those elsewhere in the world, questions arise as to the extent and uniqueness of the Antarctic haloarchaeal pan-genome.
In a study just published in Microbiome we set about defining differences between Hrr. lacusprofundi strain R1S1 (from Rauer 1 Lake) and strain ACAM34 (from Deep Lake). The strains provided the capacity to compare isolates from lakes located in different sectors of the East Antarctica region (one of 16 distinct ice-free biogeographic regions of Antarctica – Terrauds and Lee Diversity Distrib 2016): Rauer 1 Lake from Filla Island in the Rauer Island group, and Deep Lake ~ 30 km away and ~9 km ENE from Davis Station in the Vestfold Hills (image 1).
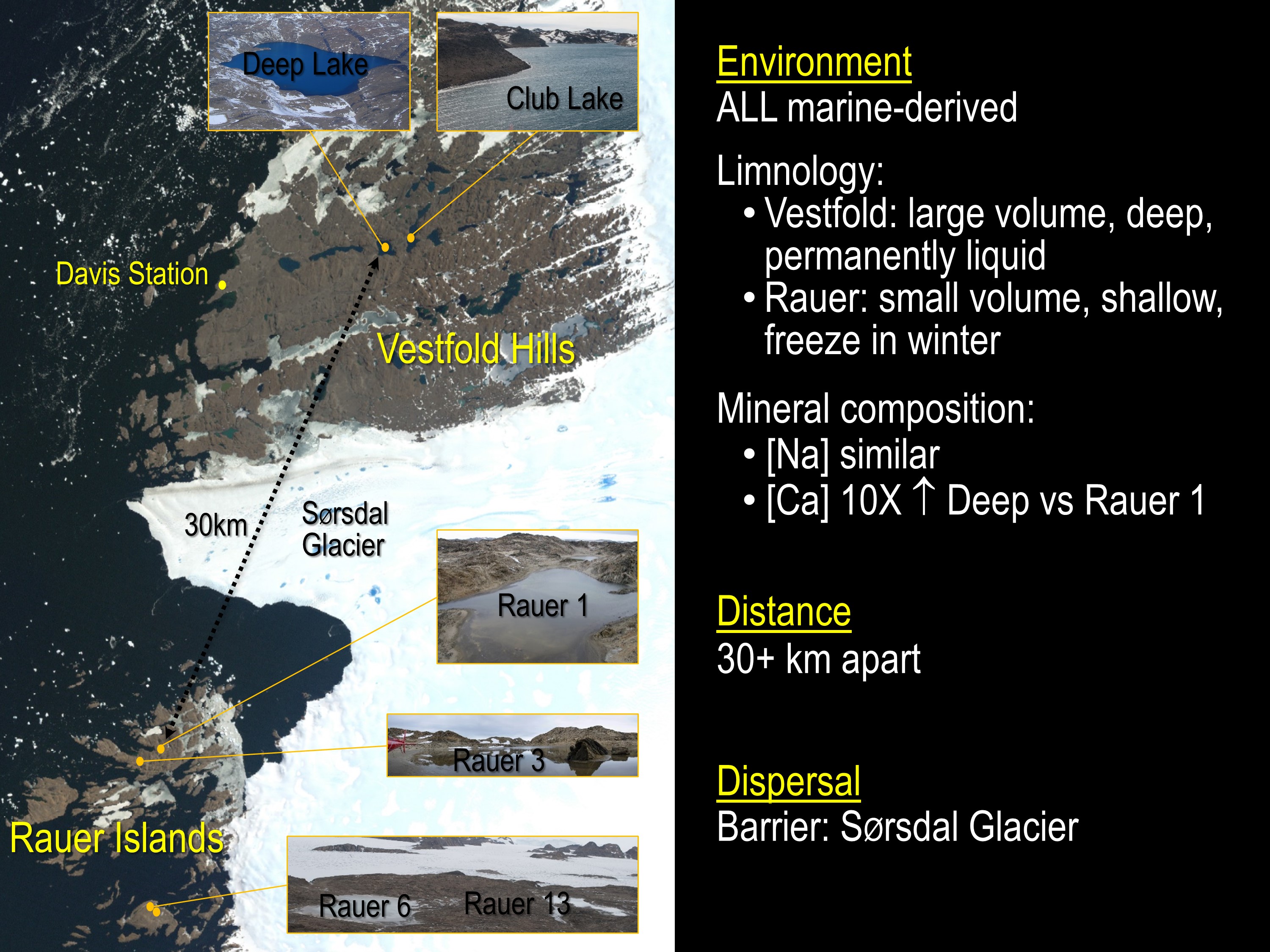
The analysis of strain variation formed the last chapter of Bernhard Tschitschko’s PhD studies. His analysis coincided with data beginning to churn out from our JGI Community Science Program that was (and still is) producing metagenome data from samples taken during a major 2013-2015 over-wintering expedition along with some others samples from 2006 and 2008. As a result, Bernhard and others (from my group, JGI and UTS) were able to analyse metagenome data generated from four hypersaline Rauer Island lakes (Rauer 1, 3, 6 and 13), Club Lake (which neighbours Deep Lake) and a Deep Lake time series (2006, 2008, 2013-2014). This enabled assessments of population level genomic variation including the prevalence of strain specific genomic markers, and biogeographic patterns of genome evolution.
Because the work involved a very detailed analysis of genomic content and variation through to a comparative assessment of communities across disparate lake systems, the types of outcomes realized from the study were quite varied. For example, Bernhard found that the way that haloarchaea accommodate genomic variation relates to replicon structuring (image 2). Different species of haloarchaea have one replicon (chromosome) or multiple replicons (one large primary replicon and one or more secondary replicons). Bernhard determined that haloarchaea which have a high proportion of their genome contained in secondary replicons have a proportionately lower content of genomic variation in their primary replicon (and vice versa).

Consistent with Bernhard's previous study (Tschitschko et al ISMEJ 2015), he found that most of the Hrr. lacusprofundi strain variation could be associated with host-virus interactions (evasion/defence), with an interesting feature being the presence of an entire CRISPR/Cas region (~9.4 kb) representing a shared HIR with DL1 (a low abundance Halobacterium species). The study was also the first to use an Antarctic virus (isolated by Suzanne Erdmann) to perform infection studies on the two strains of Hrr. lacusprofundi, determining that one strain was easily infected and lysed, while the other strain was resistant.
In terms of grander implications, the study showed that specific haloarchaeal species were major species across the Rauer Islands and Vestfold Hills hypersaline lake systems (image 3).
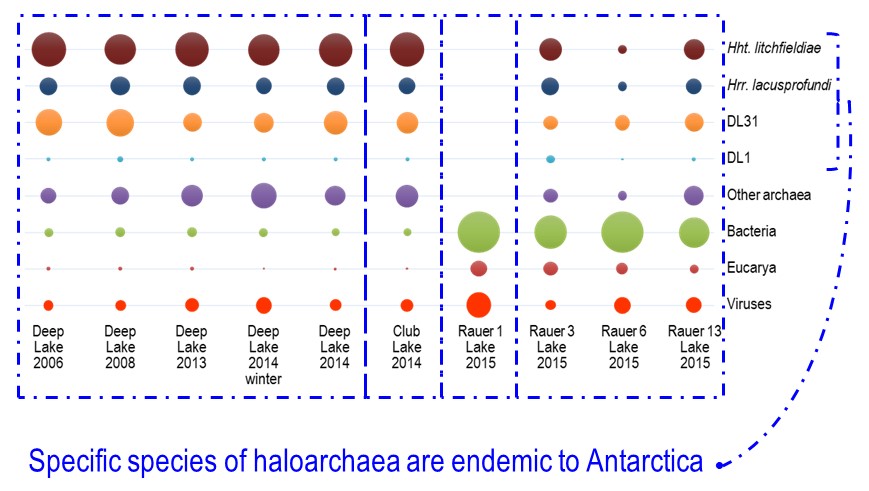
The dominant haloarchaeal populations possessed a high level of genomic conservation and also exhibited biogeographic distinctions (image 1 & 4).
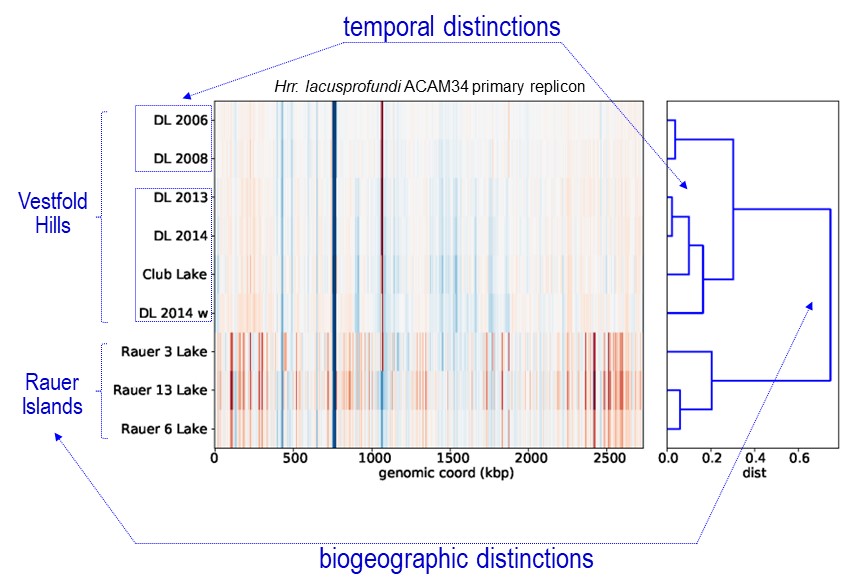
This indicates that these species are overall endemic to the Vestfold Hills-Rauer Islands and may very well be endemic to Antarctica. Very little is known about the endemism of Antarctic microorganisms (Cavicchioli Nat Rev Microbiol 2015): do regional distinctions occur (i.e. between the 16 distinct ice-free regions)?; does Antarctica have its own ‘total microbial pan-genome’ that varies to the rest of the world?; what does the extent of endemism mean for species invasion? – will the Antarctic populations be susceptible to alien species and hence ecosystem function become irrevocably perturbed? – one of our earlier studies suggests this could be the case for viral invasion of keystone bacterial species in Ace Lake (Lauro et al ISMEJ 2011).
Antarctic wildlife has some very real and profound issues to deal with. Antarctic waters are already experiencing pollution (plastics, drugs, wastes) and a high risk of invasive species and diseases due to several million people visiting per year (tourism, fisheries, research stations). In a focus issue of Nature on Antarctica, it was highlighted that Antarctica has in fact lost its ‘pristine’ status due to untoward human behaviour (Nature Editorial 2018). Hopefully it is studies like ours that will help to document just how much natural wonder exists and waits to be discovered in Antarctica – there are all the right reasons in the world to motivate sensible politics towards providing real and lasting protection for this unique, polar environment (Cavicchioli Greenpeace).
Article poster image photo credits: Alyce Hancock, Sarah Payne, Rick Cavicchioli
Follow the Topic
-
Microbiome
This journal hopes to integrate researchers with common scientific objectives across a broad cross-section of sub-disciplines within microbial ecology. It covers studies of microbiomes colonizing humans, animals, plants or the environment, both built and natural or manipulated, as in agriculture.

Related Collections
With Collections, you can get published faster and increase your visibility.
Oncobiome
This collection of papers delves into the burgeoning field of oncobiome research, exploring the intricate relationship between cancer and the microbiome. The oncobiome encompasses the diverse microbial communities residing in and on the human body, which influence cancer development, progression, and treatment responses. By examining these interactions, our aim is to unravel the complex mechanisms through which the microbiome impacts oncogenesis and therapeutic outcomes.
This compilation highlights cutting-edge research, offering insights into potential diagnostic markers and novel therapeutic strategies, thereby advancing our understanding of cancer biology and paving the way for innovative, microbiome-targeted cancer treatments.
This is a cross-journal collection between:
Experimental Hematology and Oncology
Articles will undergo the standard peer-review process of the journal to which they are submitted and are subject to either the BMC editorial policies or those of BJC Reports. Articles will be added to the Collection as they are published. The Editors have no competing interests with the submissions which they handle through the peer review process. The peer review of any submissions for which the Editors have competing interests is handled by another Editorial Board Member who has no competing interests.
Publishing Model: Open Access
Deadline: Ongoing
Animal Gut Nutrition and Greenhouse Gas Mitigation
Animal Microbiome, Journal of Animal Science and Biotechnology and Microbiome call for submissions to the collection on Animal Gut Nutrition and Greenhouse Gas Mitigation.
Efforts to reduce greenhouse gas emissions from livestock systems increasingly hinge on innovations in animal gut nutrition. The dynamic relationship between the gut microbiome and nutrient utilization plays a pivotal role in shaping methane output, feed efficiency, and overall sustainability. Advances in microbial ecology—particularly in understanding the role of gut microbiome in nutrient metabolism—are opening new pathways for mitigating emissions while enhancing productivity. These developments support the implementation of climate-smart agricultural strategies to address climate change and its impacts.
Looking ahead, continued research in this field has the potential to yield innovative solutions such as targeted probiotic supplementation, which could further optimize gut function and enhance nutrient absorption. These advancements may lead to reduced greenhouse gas emissions while improving animal health and productivity. By deepening our understanding of the animal gut microbiome, we can contribute significantly to sustainable agricultural practices that benefit both the environment and food security.
We invite researchers to contribute to this special Collection on Animal Gut Nutrition and Greenhouse Gas Mitigation. Topics of interest include but are not limited to:
- Animal Gut Microbiome and Feed Efficiency
- Greenhouse Gas Mitigation Strategies
- Rumen Fermentation Dynamics
- Nutrient Utilization in Livestock
- Probiotic Supplementation Effects
- Sustainable Livestock Production Practices
- Climate-Smart Agriculture Innovations
This Collection supports and amplifies research related to SDG 13, Climate action.
All submissions in this collection undergo the relevant journal’s standard peer review process. Similarly, all manuscripts authored by a Guest Editor(s) will be handled by the Editor-in-Chief of the relevant journal. As an open access publication, participating journals levy an article processing fee (Animal Microbiome fees, Journal of Animal Science and Biotechnology fees, Microbiome fees). We recognize that many key stakeholders may not have access to such resources and are committed to supporting participation in this issue wherever resources are a barrier. For more information about what support may be available, please visit OA funding and support, or email OAfundingpolicy@springernature.com or the Editor-in-Chief of the journal where the article is being submitted.
Publishing Model: Open Access
Deadline: Sep 04, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in