Cyclic di-GMP is central to the function of T3SSs
Published in Microbiology
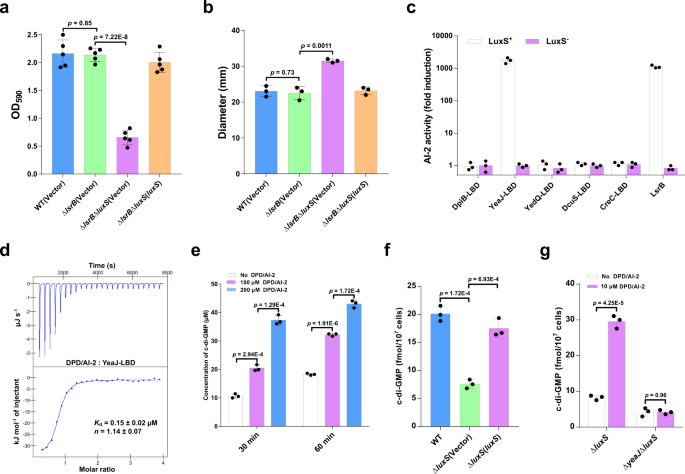
While playing important functions, bacterial secretion systems such as type III secretion systems (T3SSs), type IV secretion systems (T4SSs) and type VI secretion systems (T6SSs) require energy transduction systems to power the export of their substrates through the outer membrane. To avoid unnecessary energy consumption, bacteria turn these protein secretion machines on when required and turn them off when their activities are not needed. Bacterial secretion systems are regulated precisely by a variety of regulatory systems at transcriptional, translational, and post-translational levels, which enable bacteria to adapt to changing environments. Remarkably, cyclic di-GMP (c-di-GMP) signaling has been shown to regulate T3SSs, T4SSs and T6SSs in pathogenic bacteria such as Salmonella enterica serovar Typhimurium, Pseudomonas aeruginosa, and Agrobacterium tumefaciens1-3. However, extracellular signals that modulate c-di-GMP metabolism to control bacterial secretion systems and c-di-GMP effectors that relay environmental stimuli to changes in activity of the secretion systems remain largely obscure.
S. Typhimurium is a facultative anaerobic bacterium that causes gastroenteritis in human and a typhoid-like disease in mice. S. Typhimurium encodes 17 predicted c-di-GMP-metabolizing enzymes, among which 9 contain a putative periplasmic sensory domain flanked by two transmembrane helices4. It has been reported that the DGC YedQ increases c-di-GMP concentrations through sensing of L-arginine and other nutrient-related signals by its periplasmic sensory domain4. However, it remains unclear if and how the activity of other 8 c-di-GMP-metabolizing enzymes with periplasmic sensory domains is modulated by extracellular signals. Our study reveals that the quorum sensing signal autoinducer-2 (AI-2) and bile salts induce c-di-GMP synthesis via directly targeting a GAPES1 domain-containing diguanylate cyclase (DGC) YeaJ and the transmembrane DGC YedQ, respectively. Widespread occurrence of GAPES1-containing AI-2 receptors in bacterial species belonging to Enterobacterales suggests new signal pathways for AI-2-mediated interspecies communications within the gut microbial communities. Identification of YedQ as a bile receptor in enteric pathogens including S. Typhimurium and enterohemorrhagic Escherichia coli (EHEC) suggests bile salts will induce a rise in intracellular c-di-GMP levels when these pathogens enter the host gut.
T3SS-1 encoded by Salmonella pathogenicity island 1 (SPI-1) mediates invasion of intestinal epithelial cells5. T3SS-1 has been shown to be regulated by c-di-GMP signaling, with decreased secretion of T3SS-1 effectors and reduced invasion caused by elevated c-di-GMP levels1,6. Bile salts have been shown to repress SPI-1 gene expression through unrecognized mechanisms7. Our study resolves this conundrum by linking the bile stimulus to changes in c-di-GMP concentration in S. Typhimurium. As AI-2 induces c-di-GMP synthesis, it is not surprising that AI-2 also represses T3SS-1 gene expression.
The last question is how c-di-GMP signaling control the T3SS-1. Combining reasonable speculation with experimental verification, we identify the T3SS-1 chaperone SicA as a previously unknown c-di-GMP effector and thus reveal the underlying mechamism of c-di-GMP-mediated control of the T3SS-1. Strikingly, We also find that the CesD/SycD/LcrH family of T3SS chaperones in bacterial pathogens, including P. aeruginosa, Shigella flexneri serotype 2a, Yersinia enterocolitica, EHEC O157:H7, Burkholderia thailandensis and Vibrio parahaemolyticus, are able to bind c-di-GMP, suggesting a widely conserved mechanism through which c-di-GMP signal input modulates the activity of T3SSs.
Our work provides new insights into how environmental stimuli modulate bacterial c-di-GMP metabolism, as well as the molecular mechanism underlying c-di-GMP-mediated control of bacterial T3SSs. Our study suggests a model that cell density, environmental stimuli and host-derived cues regulate c-di-GMP synthesis to control the T3SS (Figure 1). These findings will deepen the understanding of c-di-GMP metabolism and recognition in pathogenic bacteria.
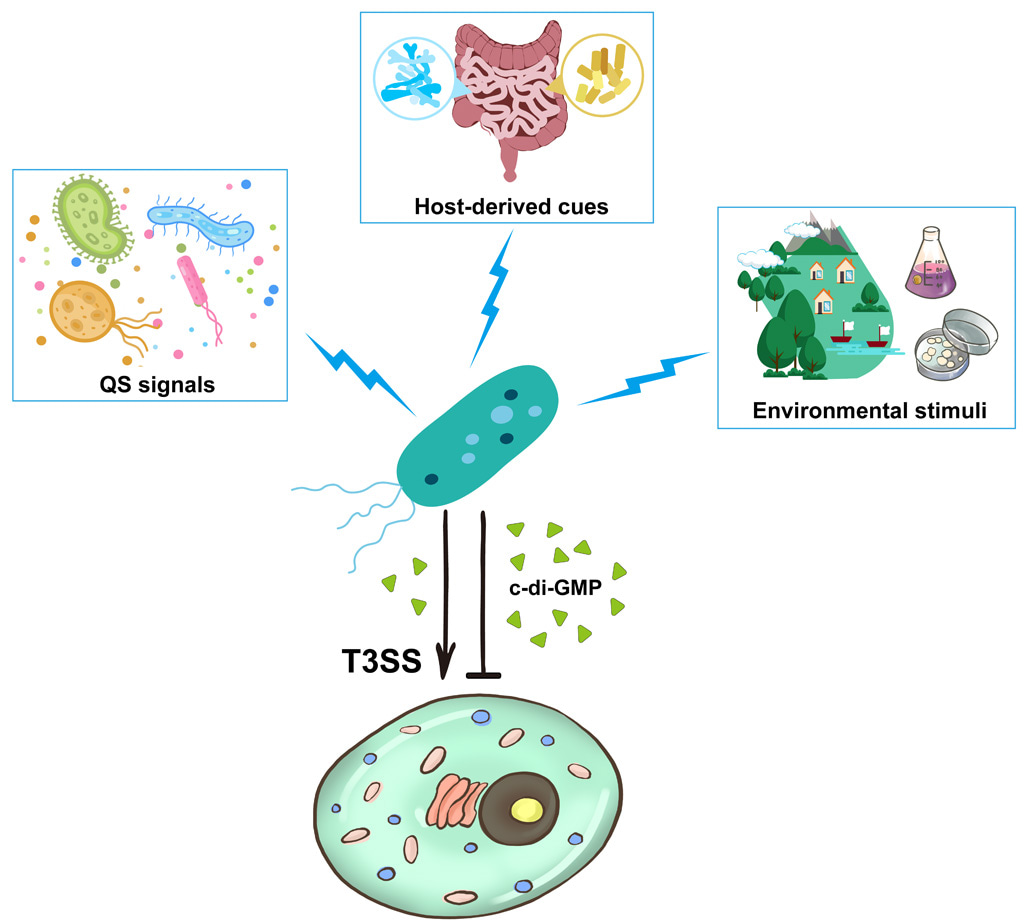
Figure 1. Conceptual diagram depicts the idea that extracellular signals modulate intracellular c-di-GMP levels to control the T3SS activity.
- Lamprokostopoulou, A., Monteiro, C., Rhen, M. &Römling, U. Cyclic di-GMP signalling controls virulence properties of Salmonella enterica serovar Typhimurium at the mucosal lining. Microbiol. 12, 40–53 (2010).
- Moscoso, J. A., Mikkelsen, H., Heeb, S., Williams, P. &Filloux, A. The Pseudomonas aeruginosa sensor RetS switches type III and type VI secretion via c-di-GMP signalling. Microbiol. 13, 3128–3138 (2011).
- McCarthy,R. R. et al. Cyclic di-GMP inactivates T6SS and T4SS activity in Agrobacterium tumefaciens. Mol. 112, 632–648 (2019).
- Mills,E., Petersen, E., Kulasekara, B. R. & Miller, S. A direct screen for c-di-GMP modulators reveals a Salmonella Typhimurium periplasmic ʟ-arginine-sensing pathway. Sci. Signal. 8, ra57 (2015).
- Hall, C. L. &Lee, V. T. Cyclic-di-GMP regulation of virulence in bacterial pathogens. Wiley Interdiscip. Rev. RNA 9, 10 (2018).
- Zheng, Y. et al. The EAL domain containing protein STM2215 (rtn) is needed during Salmonellainfection and has cyclic di-GMP phosphodiesterase activity. Microbiol. 89, 403–419 (2013).
- Prouty, A. M. &Gunn, J. S. Salmonella enterica serovar Typhimurium invasion is repressed in the presence of bile. Immun. 68, 6763–6769 (2000).
Follow the Topic
-
Nature Communications

An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.
Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Advances in neurodegenerative diseases
Publishing Model: Hybrid
Deadline: Mar 24, 2026


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in