Do living cells use elastic molecular suspensions for environment mechanosensing and a smooth ride?
Published in Cell & Molecular Biology

Do living cells use elastic molecular suspensions for environment mechanosensing and a smooth ride?
Perception of mechanical signals by living cells is important for the regulation of many cellular processes, such as cell growth, movement and differentiation, which are critical for normal cell function, and which exhibit abnormalities in the development of cancer and cardiovascular diseases. Since the discovery of the mechanosensitive behavior of cell adhesion complexes, several potential molecular mechanisms have been proposed to explain this phenomenon. Some of these mechanisms are based on the experimental observation of mechano-dependent polymerization of actin filaments by formin proteins, others on the observation of force-induced interactions between major mechanotransduction proteins localized in cell adhesion complexes, such as talin and vinculin, and still others on the stretch-dependent activation of ion channels, such as Piezo 1,2. Although all of these phenomena are important, providing many interesting insights into the biological functions of the proteins involved and highlighting potential mechanotransduction pathways that play important roles in regulating intracellular processes, there is one feature that unites them all: each of the proposed mechanisms focuses on a single pathway, without providing a clear, holistic picture of how these mechanisms work together to endow cell adhesion complexes with a mechanosensitive function that is consistent with quantitative experimental observations of their formation, maturation and mechanotransduction.
Answering such questions often requires detailed mathematical modeling, which almost always leads to the discovery of large gaps from a physical point of view in the molecular mechanisms proposed based on purely experimental studies, sometimes taking months of brainstorming to resolve. Given the overall complexity of the processes involved in the formation, shaping and mechanotransduciton of cell adhesion complexes, this makes the problem of understanding the mechanosensitive behavior of cell adhesion one of the most challenging tasks in the mechanobiology field. This is where our journey began, with the main goal of making sense of experimental observations accumulated to date.
With such a long road ahead, it always hard for newcomers to join a large research field with so many published experimental and theoretical studies, as the saying goes, the first step is often the hardest. For this reason, we started with the simplest thing – we took an already published theoretical model accepted by the research community and turned it around this way and that to see if it can be tweaked, adding something new to improve its functionality. The most obvious choice was the so-called molecular clutch theory, which proposes that cellular mechanosensitivity is mediated by network of interacting proteins, such as talin and integrin, that physically link the actin cytoskeleton to the substrate, forming so-called “molecular clutches” that transmit the mechanical signal of the microenvironment to the cell and collectively determine the cellular response [1,2].
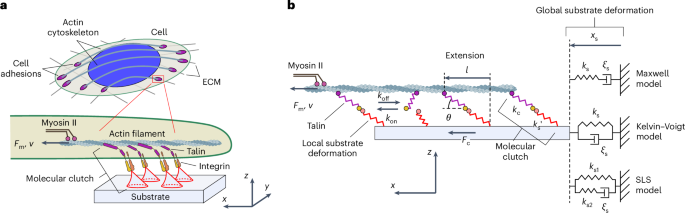
Indeed, molecular clutch theory has previously been used to qualitatively explain various aspects of cell spreading, migration and traction force generation by cells on substrates with different elasticities, providing valuable insights into these processes. However, an initial test of the model showed that it would be quite a bumpy ride – the conventional molecular clutch theory appeared to predict very erratic dynamic behavior of cell adhesion complexes, with very large fluctuations in the force they experience due to the so-called load-and-fail mechanism, as well as strong fluctuations in the retrograde flow of actin filaments that are attached to these complexes, see Figure 1(a,c). Yet, fluctuations of such large magnitude have never been observed in experiments, suggesting that while the conventional molecular clutch theory was able to provide many interesting insights into the mechanosensitive behavior of cell adhesion and migration in the past, living cells have actually come up with a smarter way of adhering to an underlying substrate and moving between two different points in space that does not require a lot of uncomfortable shaking.

Figure 1. 'Load-and-fail' mechanism versus 'differential transmission' mechanism. (a, b) Schematic representations of 'load-and-fail' and 'differential transmission' mechanisms, respectively. The formation of molecular clutches results in transmission of the force generated by myosin II motors to the substrate, causing its deformation. In the 'load-and-fail' mechanism, when the increasing tension of molecular clutches reaches a certain value, the molecular clutches dissociate in a highly cooperative manner as a result of the high rigidity of molecular clutches, which is usually used in conventional molecular clutch models, see panel (a). In contrast, in the models developed in our study, molecular clutches exhibit highly elastic properties, which is in good agreement with experiments. This leads to a loss of synchronization of the dissociation of molecular clutches under the myosin II-generated load, and as a result, the molecular clutch system exhibits a more uniform behavior, functioning as a 'differential transmission' that mechanically couples the substrate and actin cytoskeleton moving at different speeds. (c-e) Stochastic kinetic simulations of the conventional molecular clutch model and the linear WT and linear KD models developed in our study. From the plots it can be seen that while the conventional molecular clutch model predicts stochastic oscillations between zero and the maximum cell traction on soft substrates ('load-and-fail' mechanism), the linear WT and linear KD models predict much milder fluctuations of the molecular clutch system around its steady-state, with the amplitude of fluctuations being smaller in the case of softer molecular clutch components (linear WT model) due to the 'differential transmission' mechanism schematically shown in panel (b).
Quick screening of equations describing the dynamics of cell adhesion complexes revealed that to suppress unwanted vibrations, cells likely use molecular suspensions consisting of two parts: 1) highly elastic adaptor proteins (e.g., talin and vinculin) that connect the actin cytoskeleton to adhesion receptors, and 2) a substrate locally deformed at the adhesion sites of individual molecular clutches, providing additional elasticity to the force-transmitting molecular clutches. Calculations have shown that in sharp contrast to the conventional load-and-fail mechanism, which requires actin filaments to flow and the underlying substrate to deform at nearly synchronous rates to the point of complete failure of all molecular clutches connecting them, repeating the cycle over and over [Figure 1(a,c)], the modified model predicts that elastic composite molecular suspensions allow retrograde actin flow and substrate deformation to occur at different rates while maintaining a stable mechanical coupling between them – via differential transmission mechanism [see Figure 1(b,d,e)].
Using this new model, we were able not only to systematically fit previously published experimental data on retrograde actin flow and traction force exerted by cells on substrate, measured as a function of substrate elasticity and viscosity [2,3], and to describe the force-loading rates of molecular clutches and the effect of vinculin on it [4,5], but also to accurately depict the initial stages of nascent adhesion growth leading to their maturation into focal adhesion complexes. More importantly, it was found that the developed approach also provides a long-sought link between the aforementioned experimental data collected at the cellular scale and single-molecule studies of the force-dependent response of individual proteins involved in the assembly of cell adhesion complexes and cellular mechanotransduction, such as talin and vinculin [6-8].
In the near future, it will be interesting to further refine the model to include a description of the phase-separation processes that have been experimentally observed to promote the formation and growth of cell adhesion complexes, as well as force-induced reorganization of the actin cytoskeleton, in order to gain a more detailed understanding of the interplay between different mechanotransduction pathways in shaping the mechanosensitive behavior of cell adhesion complexes, which play a crucial role in many cell migration-dependent processes.
References
[1] C. E. Chan and D. J. Odde, Traction dynamics of filopodia on compliant substrates, Science 322, 1687 (2008).
[2] A. Elosegui-Artola, R. Oria, Y. Chen, A. Kosmalska, C. Perez-Gonzalez, N. Castro, C. Zhu, X. Trepat, and P. Roca-Cusachs, Mechanical regulation of a molecular clutch defines force transmission and transduction in response to matrix rigidity, Nat. Cell Biol. 18, 540 (2016).
[3] Mark Bennett, Marco Cantini, Julien Reboud, Jonathan M. Cooper, Pere Roca-Cusachs, and Manuel Salmeron-Sanchez, Molecular clutch drives cell response to surface viscosity, PNAS, 115:1192-1197 (2018).
[4] M. H. Jo, P. Meneses, O. Yang, C. C. Carcamo, S. Pangeni, and T. Ha, Determination of single-molecule loading rate during mechanotransduction in cell adhesion, Science 383, 1374 (2024).
[5] Y. Hu, H. Li, C. Zhang, J. Feng, W. Wang, W. Chen, M. Yu, X. Liu, X. Zhang, and Z. Liu, DNA-based ForceChrono probes for deciphering single-molecule force dynamics in living cells, Cell 187, 3445 (2024).
[6] M. Yao, B. T. Goult, H. Chen, P. Cong, M. P. Sheetz, and J. Yan, Mechanical activation of vinculin binding to talin locks talin in an unfolded conformation, Sci. Rep. 4, 4610 (2014).
[7] M. Yao, B. T. Goult, B. Klapholz, X. Hu, C. P. Toseland, Y. Guo, P. Cong, M. P. Sheetz, and J. Yan, The mechanical response of talin, Nat. Commun. 7, 11966 (2016).
[8] Y. Wang, M. Yao, K. B. Baker, R. E. Gough, S. Le, B. T. Goult, and J. Yan, Force-dependent interactions between talin and full-length vinculin, J. Am. Chem. Soc. 143, 14726 (2021).
Follow the Topic
-
Nature Physics
This journal publishes papers of the highest quality and significance in all areas of physics, pure and applied.


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in