Group formation is a social affair
Published in Microbiology

The first steps toward biological complexity
Most forms of life on our planet exhibit remarkable complexity due to the interplay of various cell types, all performing highly specialized functions. Like the gears in a clock, the cells in these animals or plants work in accordance to result in complex functions such as sight, root development, and - in the case of the clock – keeping time. The first evolutionary step towards such complexity is the formation of multicellular organisms1 rather than remaining independent unicellular organisms. Multicellularity has evolved multiple times in the evolution of life on Earth. It is commonly accepted to be preceded by the formation of temporary groups of cells that can form and disband back to their unicellular mode of life. At the scale of microorganisms, such temporary groups can offer various benefits to the constituent cells, such as protection against predators and chemicals2. While these benefits can often signify the difference between life and death, forming and maintaining multicellular groups comes at a metabolic cost, for example, the production of compounds that can keep cells together. However, ‘cheater’ cells that do not produce these compounds can exploit ‘cooperator’ cells and use their compounds to incorporate themselves into the groups3. As they do not carry the cost of production, cheaters will grow faster than the cooperators and eventually drive them to extinction, consequently losing the ability to form groups and stumping further evolution of multicellularity4.

of unicellular organisms. Adapted from Fisher & Regenberg.2
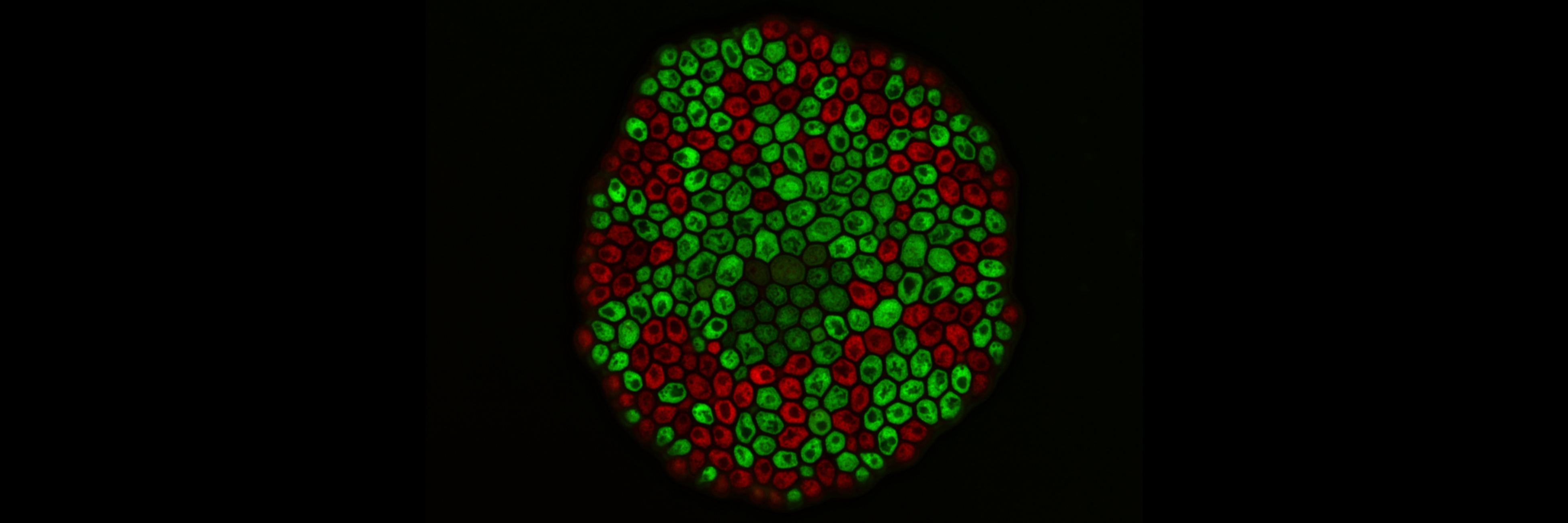
Yeast has a sticky green beard
In our work, we study the group formation of baker’s yeast by a process called flocculation. In flocculation, yeast cells aggregate through the expression of the FLO1 gene, which encodes for a protein on the cell wall that can bind to sugar residues on the cell wall of neighboring cells. As such, expression of FLO1 allows cells to stick to - and form groups with – touching cells5. By measuring the strength of adhesion between FLO1-expressing ‘cooperator’ cells and non-expressing ‘cheater’ cells, we found that interaction between two cooperators is approximately twice as sticky as the interaction between a cooperator and a cheater. Whereas this increase in adhesion due to the potential of forming two-way bonds might seem logical from a physical perspective, this has two far-reaching ecological implications for group formation. First, since cooperators more strongly interact with each other, they could preferentially retain other cooperators in the groups. As such, the discrepancy in interactions allows the cooperators to ‘recognize’ other cooperators and steer the benefits of group formation towards them. As such, the FLO1 gene can be considered to be a so-called ‘green beard’ gene, as it governs both the cooperative benefits and kin recognition towards other cooperators. Second, as the interactions with a non-expressing cell are nonzero, the kin recognition mechanism is imperfect and these cells can still be incorporated into the groups.
At the mercy of flow
To be able to group up, yeast cells have to bump into other yeast cells. However as Saccharomyces lacks an intrinsic motility mechanism, yeast cells are subject to environmental flows to collide with each other. Similarly, formed groups can also be disrupted by strong environmental flows. To study the effects of shear flow on the aggregation of yeast cells through FLO1, we use an individual-based model where a mix of cooperator and cheater cells can aggregate in a simple shear flow. We observe that in weak flows, yeast can easily aggregate and form large groups of cells. However, cheater cells are easily incorporated into these groups as the disruptive force in these flows is not strong enough to remove cheaters from the groups. In contrast, strong flow reduces the group sizes but is relatively selective for cooperators. As such, environmental flow can modulate the size of the groups at the expense of selectivity and vice versa.

Flexibility vs. Complexity?
The social dynamics between cheaters and cooperators are not only determined by cluster size and composition, but also by the cost of expressing FLO1. After the initial flocculation step, cells residing in larger groups get higher chances of surviving an external threat, for example, a toxic chemical. All cells that survive are allowed to grow and divide taking into account a slower growth rate for the cooperators. We can determine how the frequency of cooperators and cheaters in the population would evolve by cycling these three steps; flocculation, selection, and growth. If we impose very weak selection, meaning that small groups or even single cells would survive, cooperators will eventually become extinct and group formation will be lost. Similarly, if there are many generations of growth between flocculation events, the defectors will overtake the population due to the slower growth of the cooperators. Even if only the largest groups survive, sufficient generations in between the flocculation events will result in loss of cooperation and hence loss of group formation.

A large range of flow, selection, and growth conditions result in the stable coexistence of cheaters and cooperators in the population. As such, the formed groups are always combinations of both cell types, where the cheaters are invested in their individual gain rather than the performance of the group. This consistent internal conflict between cooperators and cheaters in aggregative groups likely opposes the further evolution of complex multicellularity. On the other hand, the continued presence of both cell types allows some flexibility of the population to various conditions. Cooperators can govern group formation if there is a need for its protective benefits. If there is no need, cheaters can speed up population growth due to their more efficient growth. Additionally, as flocculation permits the coexistence of different cell types, even those not invested in aggregative group formation, it can potentially conserve other group-forming mechanisms. Whether or not aggregative group formation assists or halts eventual multicellularity remains to be seen.
Discover the paper here:
References:
- Szathmáry, E. Toward major evolutionary transitions theory 2.0. Proc. Natl. Acad. Sci. U. S. A. 112, 10104–10111 (2015).
- Fisher, R. M. & Regenberg, B. Multicellular group formation in Saccharomyces cerevisiae. Proc. R. Soc. B Biol. Sci. 286, (2019).
- Smukalla, S. et al. FLO1 is a Variable Green Beard Gene that Drives Biofilm-like Cooperation in Budding Yeast. Cell 135, 726–737 (2008).
- Pentz, J. T. et al. Ecological Advantages and Evolutionary Limitations of Aggregative Multicellular Development. Curr. Biol. 30, 4155-4164.e6 (2020).
- El-Kirat-Chatel, S. et al. Forces in yeast flocculation. Nanoscale 7, 1760–1767 (2015).



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in