Harnessing directed evolution to improve plastic degrading enzymes
Published in Chemistry


Plastics serve an important role in modern day life. With a wide range of formulations designed to serve a variety of functions, plastics have become integral to innovation in many sectors such as construction, food and health.1 Unfortunately, the inherent durability of these manmade polymers means that they become persistent pollutants if disposed of incorrectly: their environmental breakdown is incredibly slow spanning hundreds, if not thousands, of years.2 The scale of this problem has led to an urgent search for new methodologies for recycling plastic waste. One particularly interesting option is to use biocatalysts to depolymerise plastic polymers into their component monomer parts.3 This idea has recently gained traction due to the discovery of an enzyme from Ideonella sakaiensis, IsPETase, which can deconstruct one of the most abundant polyester plastics, poly(ethelyne terephthalate) into the smaller chemical motifs of MHET and TPA.4
Fuelled by the discovery of IsPETase, there have been a number of efforts to tailor biocatalysts for industrial scale enzymatic PET recycling.5 Unfortunately, IsPETase in its wildtype form has limited activity and is thermally unstable, prohibiting its direct use in a commercial process. Indeed, to access the maximal rates of catalysis, you would ideally run reactions at the glass transition temperature of PET (Tg=60-70°C), where the polymer chains become more accessible to enzyme action. At these temperatures IsPETase is completely inactive. These challenges have led other researchers to focus on thermostable promiscuous homologous cutinases for PET deconstruction,6 but the question still remained as to whether the naturally evolved IsPETase could have a superior ability to depolymerise PET if only it were more thermostable.
The gold standard for improving biocatalysts for industrial use is directed evolution.7 Directed evolution (DE) is a mimic of Darwinian evolution, but on a laboratory scale (Fig. 1). A library of mutant enzymes is created and then screened for activity, with researchers directly in control of the selection pressures exerted on the biocatalyst to mould it for their application of interest. Only the top performing of these enzyme mutants are selected for further characterisation, with the best biocatalyst being subjected to further evolutionary rounds. For this process to be applied, you need a high-throughput method for evaluating all the enzyme mutants created. At the start of this project, a methodology for quantifying plastic deconstruction with sufficient throughput did not exist, and so PETases had never before been subjected to this powerful technique.
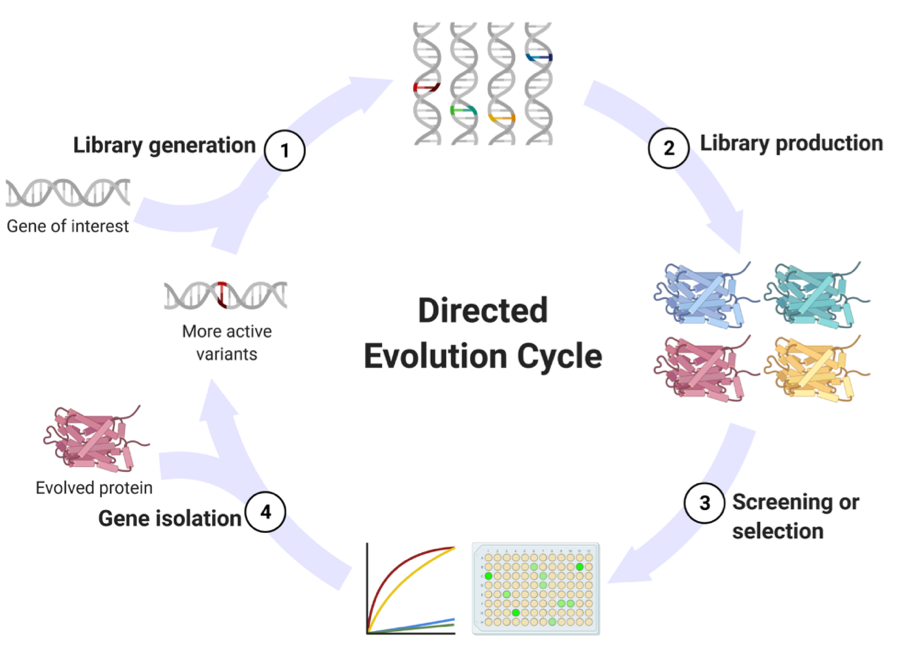
Figure 1. Directed evolution cycle. (1) First, libraries of variants are created by mutating the gene of interest. (2) The gene libraries are expressed to produce a library of proteins. (3) These proteins are then assessed for their activity either by screening for the function of interest or selection-based approaches. (4) The best protein variants are selected and the associated genes isolated. These new gene variants are then the substrate for the next round of evolution.
We aimed to design a high-throughput PET depolymerisation analysis method to enable us to subject IsPETase to directed evolution. The basic idea was to prepare a reaction with enzyme and solid PET, incubate the reaction at a desired temperature, quench the reaction and then measure the soluble products released from enzymatic plastic deconstruction, namely MHET and TPA, by HPLC (Fig. 2). Unsurprisingly, doing this in a high-throughput manner was not trivial! The first hurdle to overcome was the speed of our analysis method. The initial HPLC method for analysing the reaction products that we developed was 13 minutes long. This would take over a day to analyse the results of a single 96-well plate, and we wanted to look at at least 20 plates per evolution round. This method was therefore optimised, giving us the ability to analyse a single enzymatic plastic deconstruction reaction in just over two minutes, with a whole plate being analysed in just over three hours.
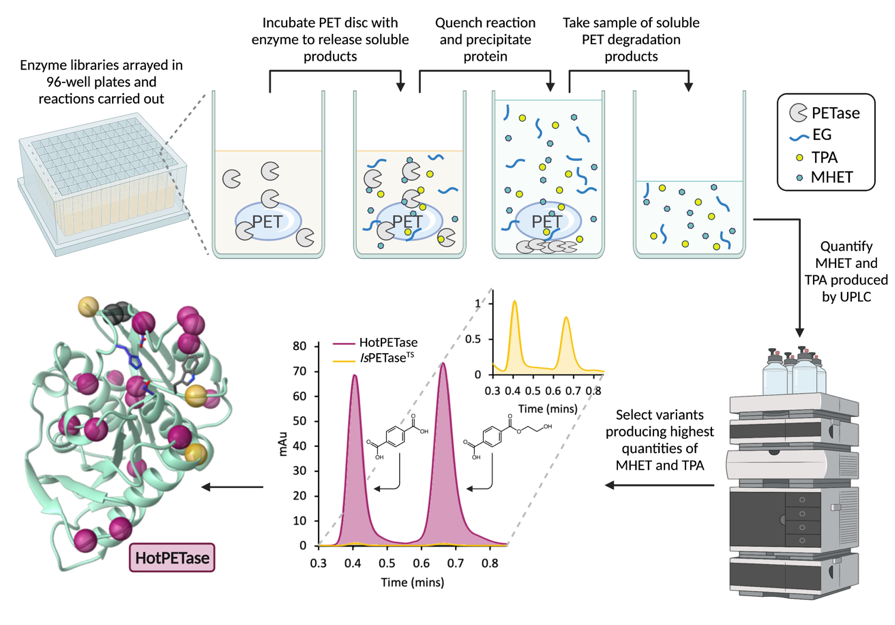 Figure 2. Workflow for the directed evolution of a PETase. Depiction of the laboratory evolution workflow for a single well in a 96-well plate. The example UPLC trace demonstrates a 24 hr PET degradation reaction using the best variant following evolution, HotPETase (pink), at 70°C as compared to the starting protein (yellow). The crystal structure shows the 21 amino acid positions mutated from IsPETaseWT.
Figure 2. Workflow for the directed evolution of a PETase. Depiction of the laboratory evolution workflow for a single well in a 96-well plate. The example UPLC trace demonstrates a 24 hr PET degradation reaction using the best variant following evolution, HotPETase (pink), at 70°C as compared to the starting protein (yellow). The crystal structure shows the 21 amino acid positions mutated from IsPETaseWT.
Next, we needed an easy method to array PET substrate across a 96-well plate. The solution was uniform discs of amorphous PET, which had to be manually hole punched from a film and painstakingly dropped into each well of the plate by hand, no small task when we were looking to screen ~2000 wells per round of evolution. The subsequent challenge was designing a screening protocol that allowed us to measure the activity of PETase mutants in cell lysate, as this would simplify our evolution workflow. This should have been fairly simple as the Green Lab is quite experienced in running lysate based assays. Individual colonies of IsPETase mutants were manually picked into 96-well plates (these were the days before we got a colony picking robot), the cells grown, pelleted and lysed, PET degradation assays set up and incubated, and then analysed by HPLC, and we saw … absolutely nothing. However, after some careful investigation it turned out that our normal cell lysis reagents, lysozyme and polymyxin B, severely hamper PET deconstruction. Hence, we had to design a new cell lysis protocol avoiding these components, by using a commercial reagent Bugbuster. Potential disaster averted we were well set to begin our evolution.
The clever part of directed evolution is that the researchers are in control of the selection pressures applied to mould an enzyme to suit your needs. With our evolution workflow set up, optimised and automated using liquid handling robots we began to engineer IsPETase by playing around with those pressures (Fig. 3). We first began to raise the reaction temperature through the rounds of evolution, weeding out any thermally labile mutants and promoting enzymes which could both withstand and react at temperatures from 60-70°C. Once we were happy with thermostability, we looked to catalyst longevity, incrementally increasing the reaction time, concentrating on selecting enzymes that could depolymerise PET at high temperatures for longer time periods. Finally, we focused on what sort of PET we wanted our enzyme to be capable of deconstructing. Amorphous PET is great as an easy substrate to array in a 96-well plate, but in reality, plastic waste actually contains a degree of crystallinity (~30%). Crystalline PET is much more difficult to depolymerise enzymatically, and so in our final rounds of DE we set our evolved IsPETase variants a new challenge, screening the best performing enzymes from our 96-well amorphous plate based assays against a crystalline PET powder. Validating the power of this approach, if we assess mutant PETases created through our evolution, we see that the proteins become increasingly more thermostable, with improved abilities to degrade crystalline PET powder over the course of 24 hours at the elevated temperature of 60°C.
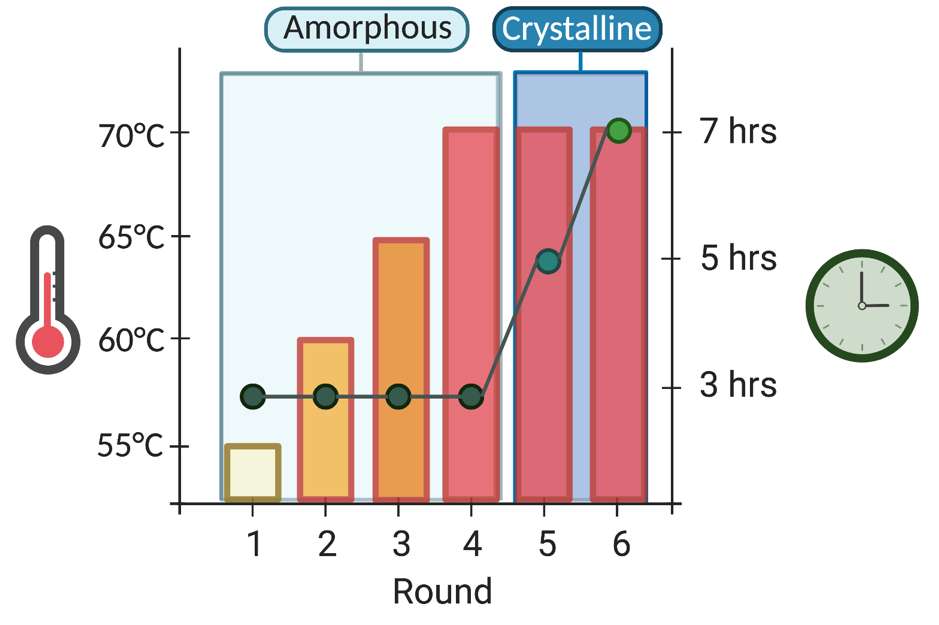 Figure 3. Selection pressures for the directed evolution of IsPETase. Across the DE rounds (x-axis), the temperature of the plastic degrading reaction was raised (left y-axis), the length of the reaction was increased (right y-axis) and the PET substrate was changed from amorphous to crystalline.
Figure 3. Selection pressures for the directed evolution of IsPETase. Across the DE rounds (x-axis), the temperature of the plastic degrading reaction was raised (left y-axis), the length of the reaction was increased (right y-axis) and the PET substrate was changed from amorphous to crystalline.
Two and a half years of evolution later, we had our final 21-point mutant, HotPETase, with a melting temperature of 82.5°C and the ability to deconstruct crystalline PET at 70°C. We were very pleased with this, but how would our enzyme compare to other PETases out there? Acquiring this information required many days of assays, with a team of three researchers dedicated to taking samples for HPLC analysis at the correct time points. At each stage the PET substrates were carefully characterised by the Shaver lab, also based at the University of Manchester. Gratifyingly, HotPETase compares really well to both the starting IsPETase variant and a promiscuous engineered cutinase LCCICCG, exhibiting rapid depolymerisation over short time periods at both low and elevated temperatures.
To rubber stamp the utility of PETases in industrial recycling, we finally wanted to demonstrate that HotPETase could do something that other recycling methods struggled with: the selective depolymerisation of PET from a real life composite plastic packaging lid. Here, we collaborated with the Haigh Lab to analyse enzymatic depolymerisation by electron microscopy. Reactions with the PET composite lids were run and samples carefully labelled and sent off to the electron microscopy team to have a closer look at what was going on. This led to some very beautiful SEM images demonstrating how PETases deconstruct PET surfaces, and confirming the selective deconstruction of PET from the composite film, leaving the PE portion untouched (Fig. 4).
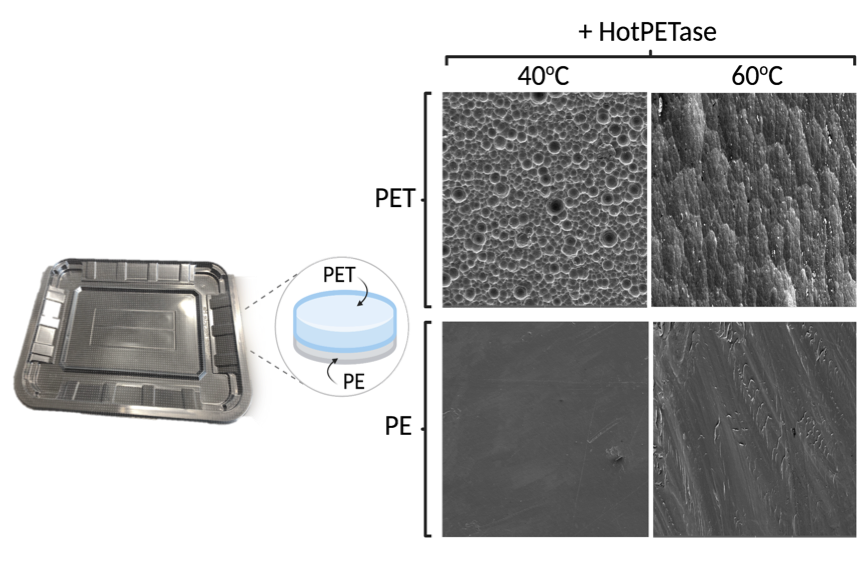 Figure 4. Biocatalytic deconstruction of a composite PET packaging lid by HotPETase. SEM images of both sides of a PET/Polyethylene (PE) packaging material degraded in the presence or absence of HotPETase, in a reaction at 40°C or 60°C over the course of six days.
Figure 4. Biocatalytic deconstruction of a composite PET packaging lid by HotPETase. SEM images of both sides of a PET/Polyethylene (PE) packaging material degraded in the presence or absence of HotPETase, in a reaction at 40°C or 60°C over the course of six days.
The icing on the cake was understanding exactly how we had changed IsPETase into a thermostable, efficient PET depolymerase. HotPETase was crystallised and characterised, with computational docking studies also carried out. The result was a nuanced understanding of how the mutations installed had imparted useful characteristics on HotPETase. Some mutations appear to aid thermostability by rigidifying secondary structures, whilst others appear to subtly change the electrostatics of the substrate binding cleft. The impact of most mutations is difficult to pinpoint, but this is the beauty of directed evolution: you do not need to directly understand how to change a protein to get your characteristic of interest, you just need to be able to identify the best performing enzymes from a pool of mutant proteins.
Through a lot of hard work, collaboration and teamwork, we present the first attempt to engineer a PET depolymerising enzyme via sequential rounds of directed evolution. Not only do we show the utility of DE for engineering multiple parameters simultaneously, but we also demonstrate that mesophilic PETases can be important and interesting starting points to begin to engineer industrially useful PET depolymerases. Most importantly, the platform we have developed can be modified to tune PETases for specific industrial reaction conditions by subtly changing the selection pressures, and could easily be deployed to develop enzymes with the ability to deconstruct other types of plastic.
We hope our research will provide a piece of the puzzle to help to solve the plastic waste problem, and hope you would like to learn more by reading our paper which can be found at: https://doi.org/10.1038/s41929-022-00821-3.
References:
- PlasticsEurope. Plastics- the Facts 2019, An analysis of European plastics production, demand and waste data. (2019).
- Geyer, R., Jambeck, J. R. & Law, K. L. Production, use, and fate of all plastics ever made. Science Advances 3, e1700782 (2017).
- Ellis, L. D. et al. Chemical and biological catalysis for plastics recycling and upcycling. Nature Catalysis 4, 539–556 (2021).
- Yoshida, S. et al. A bacterium that degrades and assimilates poly(ethylene terephthalate). Science (1979) 351, 1196–9 (2016).
- Wei, R. & Zimmermann, W. Microbial enzymes for the recycling of recalcitrant petroleum-based plastics: how far are we? Microb Biotechnol 10, 1308–1322 (2017).
- Tournier, V. et al. An engineered PET depolymerase to break down and recycle plastic bottles. Nature 580, 216–219 (2020).
- Bell, E. L. et al. Biocatalysis. Nature Reviews Methods Primers 1, 46 (2021).
Follow the Topic
-
Nature Catalysis
This journal brings together researchers from across all chemistry and related fields, publishing work on homogeneous catalysis, heterogeneous catalysis, and biocatalysts, incorporating both fundamental and applied studies.


![When PSMA-targeted therapy is not enough: high-risk localized prostate cancer after repeated [177Lu]Lu-PSMA radioligand therapy](/cdn-cgi/image/metadata=copyright,fit=scale-down,format=auto,quality=95,width=256,height=256/https://public-storage.zapnito.com/Ku6h7Yyp4Q0LXqRRMICCHR2v4LcOsmxMrmDPtOYuI1c)


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in