Harnessing functions of the microbiota to combat fungal pathogens
Published in Microbiology

Fusarium graminearum is a widespread pathogenic fungus that causes Fusarium head blight (FHB) in cereal crops worldwide, especially in wheat1. Between 2000 and 2018, an annual average of more than 4.5 million hectares were affected by FHB in China, or around 20% of the total planted area of wheat. This has resulted in annual production losses of more than 3.41 million tons2.
Wheat heads with Fusarium head blight symptoms on a few spikelets (left-hand panel), and the fungal fruiting body on a stubble (right-hand panel).
Fusarium head blight epidemics not only reduce global wheat production, but also contaminate grains with mycotoxins that threaten human and livestock health 2. To infect new hosts, F. graminearum must often endure a period of dormancy (winter) and produce spores at the right time to ensure the survival of the next pathogen generation. The main propagation strategy of this monocyclic pathogen is formation of ascospores in perithecia (fungal fruiting bodies), which are then ejected and spread via wind, insect, and rain dissemination3. Therefore, interfering with perithecium formation to decrease the initial inoculum seemed to be a promising strategy for FHB management. The screening for biological control agents (BCAs) and their application was evaluated in the frame of this study as a viable alternative and ecologically friendly method for perithecium suppression. Fungal structures are known to provide microhabitats for adapted bacterial populations, just like soil, plants, and animals. Members of bacterial communities that are intrinsically associated with plant-pathogenic fungi were therefore hypothesized to be a promising source of BCAs against highly destructive pathogens. We assumed that these bacteria likely possess advantages in terms of their adaptability to survive and proliferate on fungal structures.
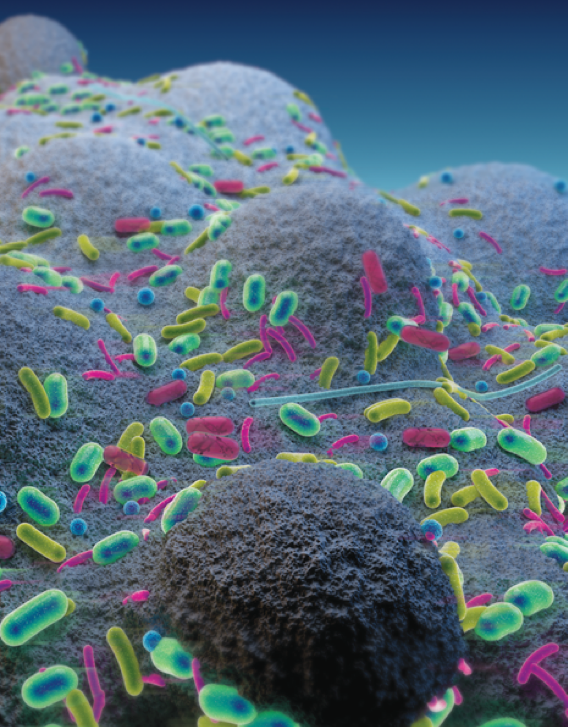
Schematic visualization of bacterial communities on the fungal perithecia
In our study, we found that F. graminearum perithecia provided a specific ecological niche for bacteria that could play an important role in disease establishment. Over 2000 bacterial strains were isolated from the native microbiota of F. graminearum, and subsequently identified and screened for antagonistic activity. We were successful in this approach and recovered 113 isolates that showed antagonistic activity against the pathogen. One specific bacterial isolate, termed as ZJU23, was identified as Pantoea agglomerans, and had the strongest inhibitory ability against F. graminearum among all isolates. Subsequently, we showed that herbicolin A secreted by ZJU23 is responsible for the observed suppression of F. graminearum.
Although herbicolin A was identified about four decades ago4,5, its biosynthetic gene cluster and mode of action against fungi were not known. In our study, we set out to investigate the biosynthetic gene cluster and mode of action against various fungi by combining various approaches, including transposon mutagenesis, liquid chromatography-mass spectrometry, atomic force microscopy and confocal microscopy. By comparing the metabolic profiles of ZJU23 and four deletion mutants of a potential biosynthesis gene cluster, we found that herbicolin A was synthesized by the AcbA-AcbJ cluster. We then proceeded to uncover the mode of action of herbicolin A against various fungi. It is important to note that the modes of action of cyclic lipopeptides against fungi are mostly unknown. What surprised us is that herbicolin A was shown to disrupt lipid rafts by interacting with ergosterol, which resulted in the formation of abnormal cell membranes, and, ultimately, caused cell death. Herbicolin A also was also found to inhibit the growth of Candida albicans and Aspergillus fumigatus, and was more effective than the clinical fungicides Amphotericin B and fluconazole. This provided important evidence for potential applications of herbicolin A beyond agriculture, for example in medicine.
Overall, we have deciphered the mechanism of the inhibitory effect of herbicolin A on fungi and identified its biosynthetic gene cluster. This could support future developments for sustainable management of Fusarium head blight worldwide.
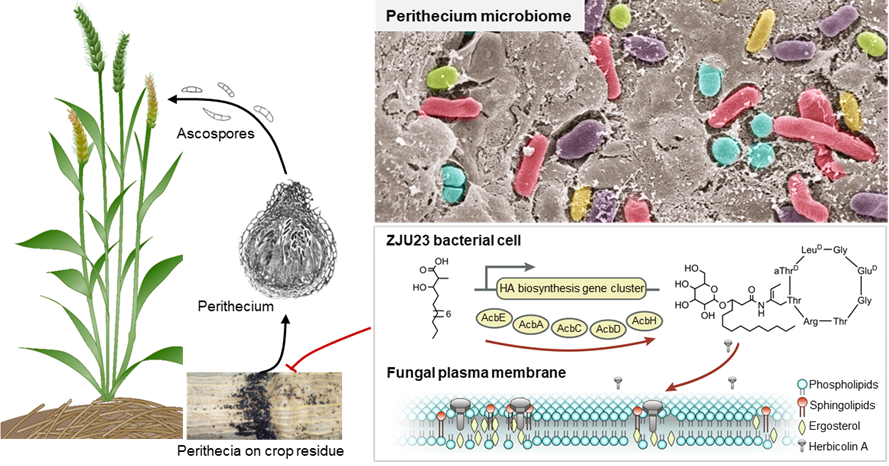
Proposed model for the mode of action of herbicolin A secreted by ZJU23 in the fungal perithecium microbiome.
Reference:
- Trail, F. For Blighted Waves of Grain: Fusarium graminearum in the Postgenomics Era. Plant Physiology 149, 103-110 (2009).
- Chen, Y., Kistler, H.C. & Ma, Z.H. Fusarium graminearum Trichothecene Mycotoxins: Biosynthesis, Regulation, and Management. Annual Review of Phytopathology, Vol 57, 2019 57, 15-39 (2019).
- Brauer, E.K., Subramaniam, R. & Harris, L.J. Regulation and Dynamics of Gene Expression During the Life Cycle of Fusarium graminearum. Phytopathology 110, 1368-1374 (2020).
- Greiner, M. & Winkelmann, G. Fermentation and isolation of herbicolin A, a peptide antibiotic produced by Erwinia herbicola strain A 111. Applied microbiology and biotechnology 34, 565-569 (1991).
- Aydin, M. et al. Structure elucidation of the peptide antibiotics herbicolin A and B. Liebigs Annalen der Chemie 1985, 2285-2300 (1985).
Yang Bai is a principle investigator at the Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, China. He received a Ph.D from University of Cologne in Germany for Arabidopsis trichome development in the lab of Professor Martin Hulskamp in 2010. From 2011 to 2016, he did a Postdoc at the Max Planck Institute for Plant Breeding Research in the lab of Professor Paul Schulze-Lefert. There, he worked on the high-throughput bacterial isolation and reconstitution of Arabidopsis root microbiota. In May 2016, he built up a research group in Beijing on the function and mechanism of plant-microbiome interactions.
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.





Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in