Imaging vesicle scission during endocytosis
Published in Protocols & Methods

Eukaryotic cells, from yeast cells to all cells in our body, probe their environment and regulate the size and composition of their surface, called the plasma membrane, by constantly engulfing tiny portions of it. This process is called endocytosis and has been involved in many essential cellular functions such as nutrient uptake, pathogen entry (among them many viruses including SARS-CoV-2), regulation of plasma membrane receptors and the formation and function of synapses in neurons and immune cells. Watching this process in living cells has thus been a major topic in cell biology since the appearance of modern live cell fluorescence imaging.
Among the many forms of endocytosis, the best characterized one is clathrin-mediated endocytosis (CME). The protein clathrin is present, together with adaptor proteins, in tiny (100-200 nm diameter) patches on the plasma membrane, forming so-called clathrin coated pits (CCPs). Clathrin remains while the plasma membrane forms an invagination and eventually a clathrin coated vesicle (CCV). After that, clathrin and associated proteins dissociate from the now vesicular membrane. The most straightforward way to watch this process in living cells is thus to mark clathrin or an associated protein with a fluorescent protein such as GFP and to take images every few seconds. Effectively, these proteins are localized in small clusters at the surface of cells. These clusters rapidly grow, remain static for a few tens of seconds, then disappear, as seen on kymograph in the Figure. These three phases of cluster life would correspond to formation of the CCP, its maturation and quick movement and/or uncoating of the CCV. This type of experiment is at the basis of most of our understanding of the mechanisms and regulation of CME [1].

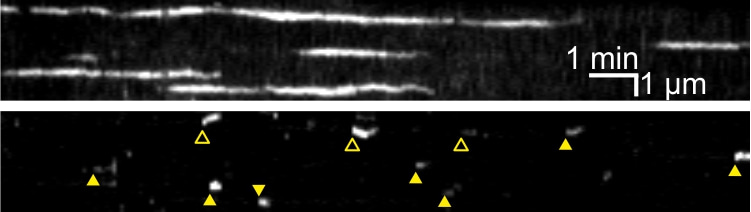
Detection of scission at clathrin coated pits. Top, kymographs of a cell co-transfected with TfR-SEP and the endocytic protein dynamin1 (Dyn1-mCherry). The top kymograph (TfR-SEP at pH 7.4) shows the appearance and disappearance of CCPs. Each CCP remains visible for more than a minute. Middle kymograph (TfR-SEP at pH5.5) shows the CCVs formed at the CCPs (yellow arrowheads). Some are associated with CCP disappearance (filled arrowheads) but for others the CCP remains visible (open arrowheads). CCV appearance is associated with the recruitment of Dyn1-mCherry a few seconds before. Bottom kymograph, the three "channels" merged. Bottom, close up view of one scission event (framed in the lila rectangle in the merge kymograph). On top, scheme of the Forming CCV with fluorescent TfR-SEP at pH 7.4 (green lollipops) and non-fluorescent TfR-SEP at pH 5.5 (black lollipops). After scission, TfR-SEP trapped in the CCV remain fluorescent when the cell is bathed in pH 5.5 solution, for the first time at frame 0 s labelled "detection". It reveals that vesicle closure occurred when extracellular pH was at 7.4, around time -2 s, "scission".
However, even if this type of measure gives a lot of information, the details of vesicular formation remain elusive. In particular, the timing of membrane scission could not be deduced from these experiments. In 2003, together with Christien Merrifield in the team of David Zenisek at Yale University, we had the idea of using another marker, the transferrin receptor (TfR), a transmembrane protein concentrated in CCPs and internalized with the CCVs. Because TfR was spanning both sides of the membrane, it could be labelled with a GFP mutant, superecliptic pHluorin (SEP), on its extracellular side. SEP is very sensitive to local pH, brightly fluorescent at neutral pH (7.4) but not at all at acidic pH (5.5). Therefore, by changing the extracellular pH from neutral to acidic, back and forth, we could determine whether the patch of TfR-SEP is still on the surface (not fluorescent during acidic perfusion) or already in a vesicle (still fluorescent despite the pH change)(see Figure). I still remember vividly when we saw how clearly the ppH protocol uncovered this process. Watching these vesicles form at the surface of living cells is still, after more than 15 years, an astonishing experience. Most importantly, it gave a very clear and temporally precise signature of vesicle formation, allowing us to describe the steps of membrane invagination and associated protein recruitment in unprecedented detail [2].
Why a Nature Protocols article 15 years after reporting this discovery? After the initial characterization and appropriate controls, we wanted to streamline the experiments and analysis. First, we designed a semi-automated detection and quantification workflow to characterize the recruitment of more than 37 associated endocytic proteins [3,4]. Last year, we finally set up a fully automated analysis workflow to get quantitative information of the endocytic activity of a living cell to test the effect of molecules interfering with CME while recording [5]. Moreover, when I present this method to various audiences, I often get questions about how to set up the synchronized fast pH changes required for the ppH protocol. I felt that the best way to answer was to write a protocol with all the details we could think of so everybody interested in this technique could more easily use it. This technique has still great potential, applied to the endocytosis of various receptors, through clathrin-dependent and independent endocytosis, in various cell types. It would also allow the localization of nascent vesicles at superresolution precision.
Finally, it was a great pleasure to write such an article with all the collaborators I had the opportunity to train in the last 10 years in my lab at Interdisciplinary Institute of Neuroscience at the University of Bordeaux. My only regret is that I could not share this with Christien Merrifield, who tragically passed away almost three years ago. This is why we have dedicated this article to his memory. Link to full text article: http://dx.doi.org/10.1038/s41596-020-0371-z
References
1. Picco, A. & Kaksonen, M. Quantitative imaging of clathrin-mediated endocytosis. Current Opinion in Cell Biology 53, 105–110 (2018).
2. Merrifield, C. J., Perrais, D. & Zenisek, D. Coupling between Clathrin-Coated-Pit Invagination, Cortactin Recruitment, and Membrane Scission Observed in Live Cells. Cell 121, 593–606 (2005).
3. Taylor, M. J., Perrais, D. & Merrifield, C. J. A High Precision Survey of the Molecular Dynamics of Mammalian Clathrin-Mediated Endocytosis. PLoS Biology 9, e1000604 (2011).
4. Cauvin, C. et al. Rab35 GTPase Triggers Switch-like Recruitment of the Lowe Syndrome Lipid Phosphatase OCRL on Newborn Endosomes. Curr. Biol. 26, 120–128 (2016).
5. Rosendale, M. et al. Functional recruitment of dynamin requires multimeric interactions for efficient endocytosis. Nat Commun 10, 4462 (2019).
Follow the Topic
-
Nature Protocols
This journal publishes secondary research articles and covers new techniques and technologies, as well as established methods, used in all fields of the biological, chemical and clinical sciences.


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in