New structures of Anion Exchanger 1 shed light on its high transport rate
Published in Healthcare & Nursing

A number of functionally important molecules and ions cannot readily diffuse through the hydrophobic plasma membranes surrounding mammalian cells. Instead, highly specialized membrane transport proteins have evolved to mediate the import and export of various otherwise poorly permeable substances. Dysfunction of these proteins impairs membrane transport which can lead to abnormal cell function and, if severe, to various whole organ disease processes.
AE1 (Anion Exchanger 1, Band 3 protein), is the predominant membrane transport protein in erythrocyte membranes and is also localized in the basolateral membrane of Type A intercalated cells in the kidney collecting duct. AE1 exchanges HCO3¯ for Cl¯ anions and this transport process is required for the normal transfer of CO2 from tissues to the lungs. In the kidney, AE1 plays an important role in urinary acidification. Naturally occurring mutations in AE1 can result in morphologic changes in erythrocytes that can lead to increased cell fragility and anemia. Mutated AE1 in Type A intercalated cells impairs urinary acidification resulting in distal renal tubular acidosis.
AE1 is one of the fastest membrane transporters known to date (~ 50,000 ions/sec), although we still do not understand how it achieves its high transport rate, which involves conformational changes of the protein and the complex coordination of ions throughout the transport pathway. This is in part due to the fact that we previously lacked knowledge of the various conformational states during the transport cycle of AE1 and the other homologous proteins that belong to the SLC4 transporter family. An X-Ray structure of the dimeric human AE1 in the outward facing open (OF) state was resolved in 2015 [1] and in previous work we used computational modelling to identify putative substrate binding sites in this state [2]. However, the OF state is just one piece of the transport puzzle of this fascinatingly rapid transporter. To understand it better, other states in its transport cycle need to be resolved – a challenging task, considering that all reported so far structures of SLC4 proteins have been resolved in the OF state. In addition, other proteins with similar architecture as SLC4 proteins (a 7+7 inverse repeat protein fold) have been resolved only in the IF state, and no 7+7-fold protein has been resolved in both IF and OF states.
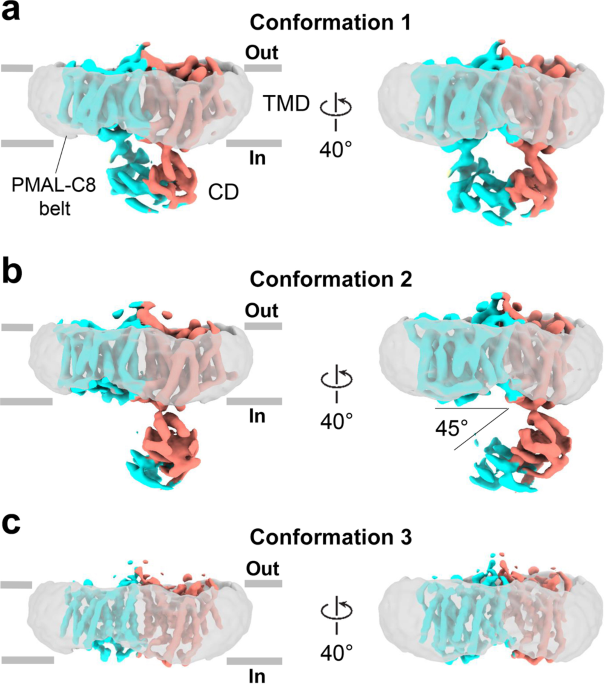
In our current work we report two new cryoEM structures of bovine AE1 (bAE1) dimers – IF-IF dimer, in which both monomers are in the IF state and a mixed IF-OF dimer, where one monomer was resolved in the OF, and the other - in the IF state. The existence of mixed dimers confirmed previous hypotheses that the two monomers of the SLC4 dimers operate independently from one another, which has been observed also in several other secondary transporters that have no structural similarity with the SLC4 proteins. Structural comparison of our new bAE1 IF and OF states, revealed that the dimeric interface (also known as a gate) is rigid and remains mostly unchanged during the IF to OF transition, while the core of the protein, which includes the substrate binding sites, rotates and shifts downward with respect to the gate. This leads to a ~5 Å vertical translocation of the amino acid residues we previously identified as a central binding site (site S1), which is likely the site where substrates bind to trigger a transport event (Fig. 1). The non-negligible vertical binding site translocation, the rigid gate domain, and the spatial separation of the core and gate domains are usually associated with an elevator transport mechanism. Thus, our new structures provide unambiguous evidence that the SLC4 family, and likely all 7+7-fold transporters, transport their substrates via an elevator mechanism as opposed to other models of transport, which have been suggested in the literature. Still, the small vertical translocation is unusual for an elevator transporter and the small structural reorganization of the protein as it morphs between the OF and IF states might hold the key to the unusually high transport rates of AE1.
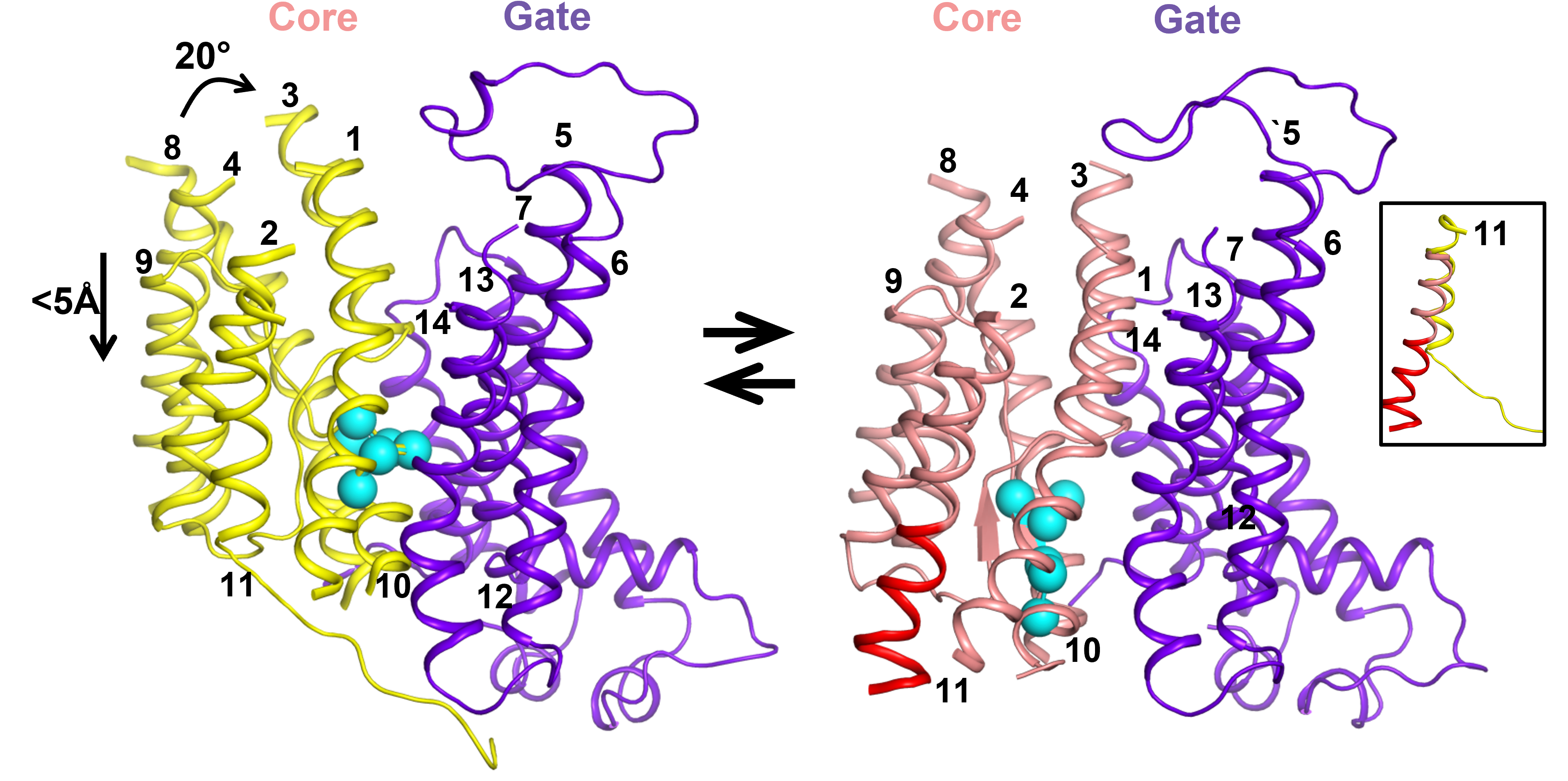
Fig.1 Elevator motion of the AE1 core between the OF state (left) and IF state (right): The core of the OF state (yellow helices) rotates and shifts downwards with respect to the rigid gate (purple helices). This results in a ~5 Å downward displacement of the residues from the central substrate binding site S1 (cyan spheres) and elongation of TM11 (red helical portion in the IF core, presented as pink helices).
Our IF AE1 structures show that transmembrane helix 11 (TM11) undergoes a large conformational change and becomes significantly longer than in the IF state. In the OF state, part of TM11 is unfolded and joins the long intracellular loop (IL5) between TMs 10 and 11 to form a β-hairpin (Fig. 2). This region of the protein therefore switches between two meta-stable states – α-helix and β-hairpin during the OF to IF transition, which may also contribute to the high rate of AE1 transport. Several mutations associated with diseases have been observed in this region and may involve impaired formation/dissociation of H-bonds, necessary for the proper structural reorganization of AE1 during the transport cycle.
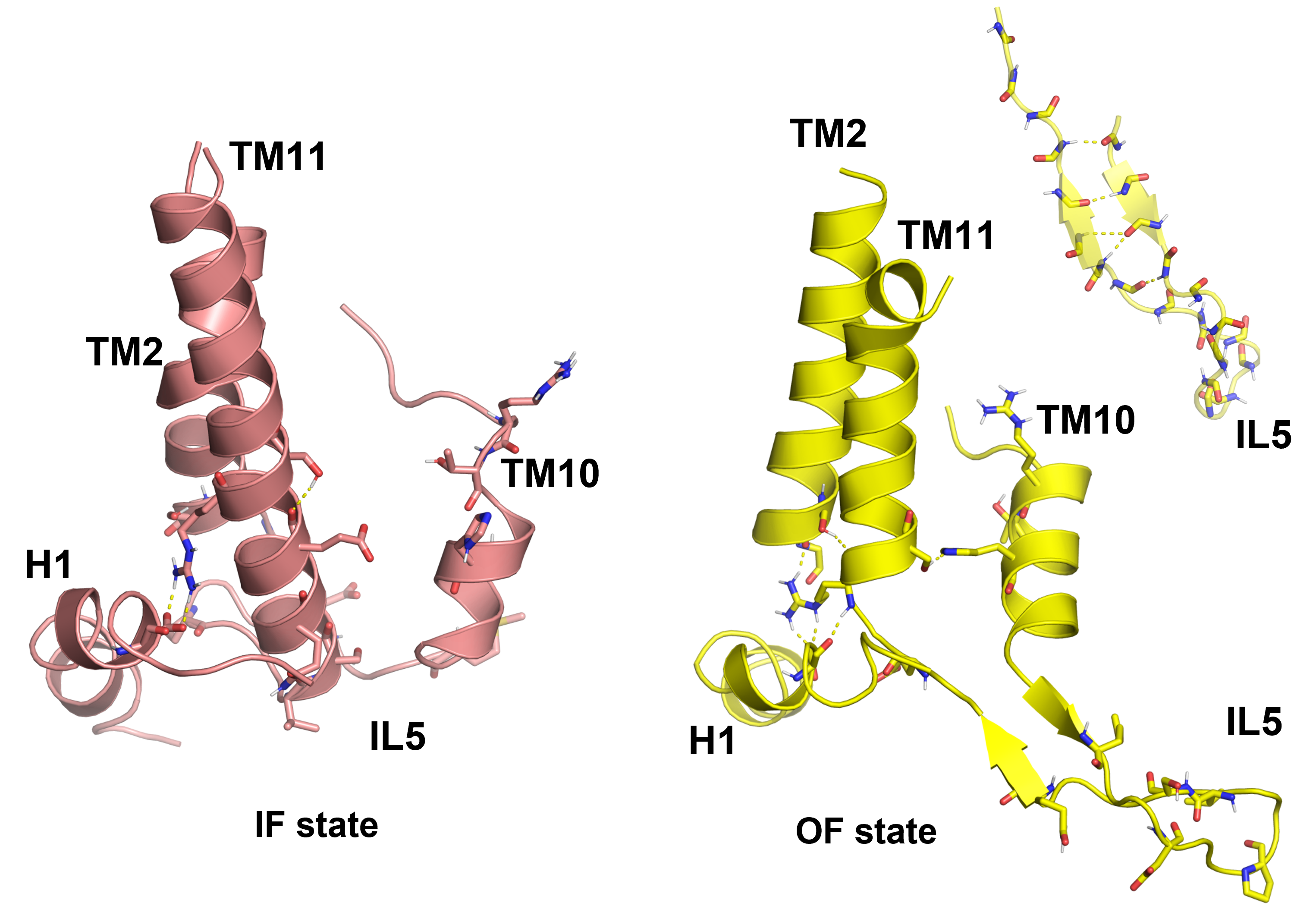
Fig.2. Structural changes in the TM10-IL5-TM11 region of AE1 in the transition from IF to OF state, during which the elongated TM11 unfolds partially and forms a β-hairpin with IL5. The residues involved in stabilizing H-bonds in this region and associated with various diseases are also shown.
To better understand the substrate dynamics in the IF and OF states of AE1 we used computational modelling to determine the distribution of ions in both states. We discovered a large reservoir for anions at the dimeric interface at the intracellular side of the protein (Fig.3), from which anions can move laterally into the IF cavity leading to the central binding site. At the extracellular side, extracellular loops prevent accumulation of anions, so the anions from the surrounding solution need to diffuse vertically through the OF cavity until they reach the central substrate binding site. Both the IF and OF cavities in the permeation pathway are large and well hydrated and we observed multiple Cl¯ and HCO3¯ entry events during 1µs long molecular dynamics simulations. The asymmetry in the ion permeation patterns may serve an important purpose. The IF state might be short-lived compared to the OF state (hence why all SLC4 structures so far have been resolved in the OF state) and is of slightly higher energy than the OF state according to coarse-grained metadynamics simulations of the IF to OF transition. Having a reservoir of anions which readily move into the IF cavity can increase the probability of anion entry and binding in the central binding site in the IF state, which then transitions into the more energetically favorable OF state. This, along with the easy substrate diffusion into the large hydrated IF and OF cavities can also contribute to the high rate of AE1 transport.
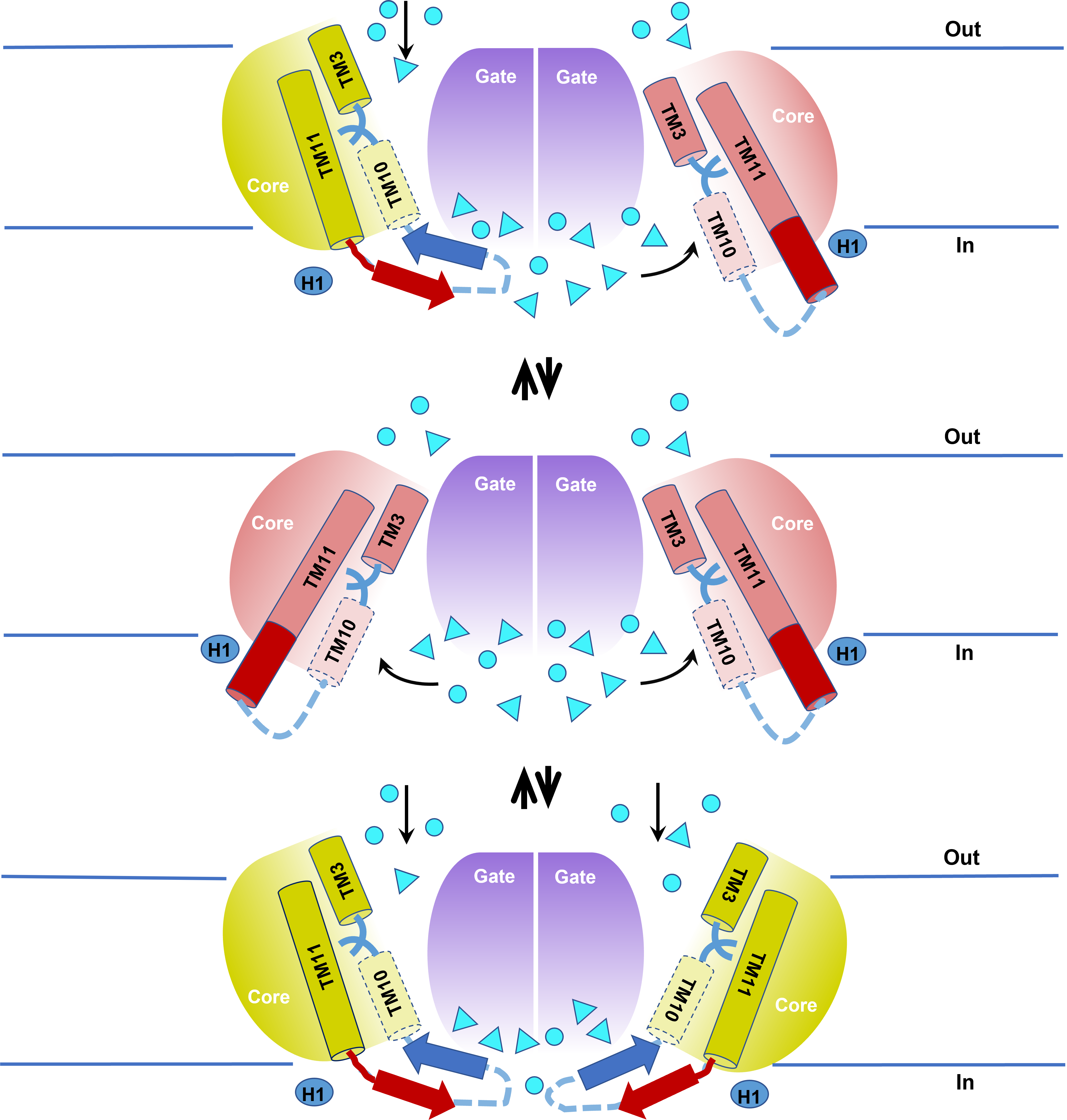
Fig.3 Scheme presenting the elevator mechanism of AE1. The Cl¯ and HCO3¯ anions are shown as cyan spheres or triangles, respectively. The figure includes some of the important findings in our study: the existence of mixed IF-OF dimers (top), the switch between a-helical (red cylinder) and b-hairpin (red and blue arrows) conformation of TM11 and IL5, the anion reservoir at the intracellular side of the dimeric gate interface and the lateral and vertical direction (black arrows) of anion entry into the IF and OF cavity, respectively.
Thus, on the basis of our new IF AE1 structures, we propose three potential reasons for the remarkable transport speed of AE1: an elevator mechanism with small structural reorganization and binding site displacement, rapid switch between two meta-stable states (α-helical and β-hairpin) of a region of the protein critical for the translocation, and swift entry and permeation of ions both in the IF and OF cavities.
References:
- Arakawa, T. et al. Crystal structure of the anion exchanger domain of human erythrocyte band 3. Science 350, 680–684 (2015).
- Wang, W. et al. Cryo-EM structure of the sodium-driven chloride/bicarbonate exchanger NDCBE. Commun. 12: 5690 (2021).
Follow the Topic
-
Communications Biology
An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.

Related Collections
With Collections, you can get published faster and increase your visibility.
Artificial Intelligence Methodology in Structural Biology
Publishing Model: Hybrid
Deadline: Nov 30, 2026
Healthy Aging
Publishing Model: Open Access
Deadline: Dec 31, 2026





Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in