Parasite life cycle innovations to invade and persist within honeybee hosts.
Published in Microbiology

Bees are keystone species in ecosystems. These insects play a major role in pollination which is essential to maintain biodiversity and contribute to crop production. Moreover, managed bees provide natural products such as honey, beeswax, bee pollen, propolis and royal jelly. In the last decades, bee health has been compromised by increasing abiotic and biotic environmental pressures leading to a reduction in their lifespan and colony fitness. Pathogen infections are one of their main threats, being trypanosomatid parasites, one of the most successful and widely distributed organisms in numerous bee species including honey bees1.
Trypanosomatid parasites (Euglenozoa: Kinetoplastea: Trypanosomatidae) are obligate eukaryotic flagellated parasites that have successfully colonized a wide range of vertebrate, invertebrate or even plant hosts. These parasites produce infections in humans but in nature they are predominantly found in insects. First evidence of trypanosomatids in honey bees date back to the early 20th century 2, with few reports later on describing their presence in the honeybee hindgut 3. To date, there are some certainties about these parasites: (i) they are commonly present in honeybees and wildbees, (ii) there are at least four species described in bees (Lotmaria passim, Crithidia mellificae, C. acantocephali and C. bombi), (iii) the flagellated unicellular state is prone to be cultured in isolation in vitro and, (iv) they could differentiate into-surface attached haptomonad stage in the ileum and rectum of their hosts. However, these are only snapshots of their life cycle that lead to other research questions such as: Are there any parasite cell innovations conferring trypanosomatids any selective advantage? Could we recapitulate the developmental program of honeybee-infective trypanosomatids parasites in vitro? Do honeybee trypanosomatid species share the same lifecycle strategies?
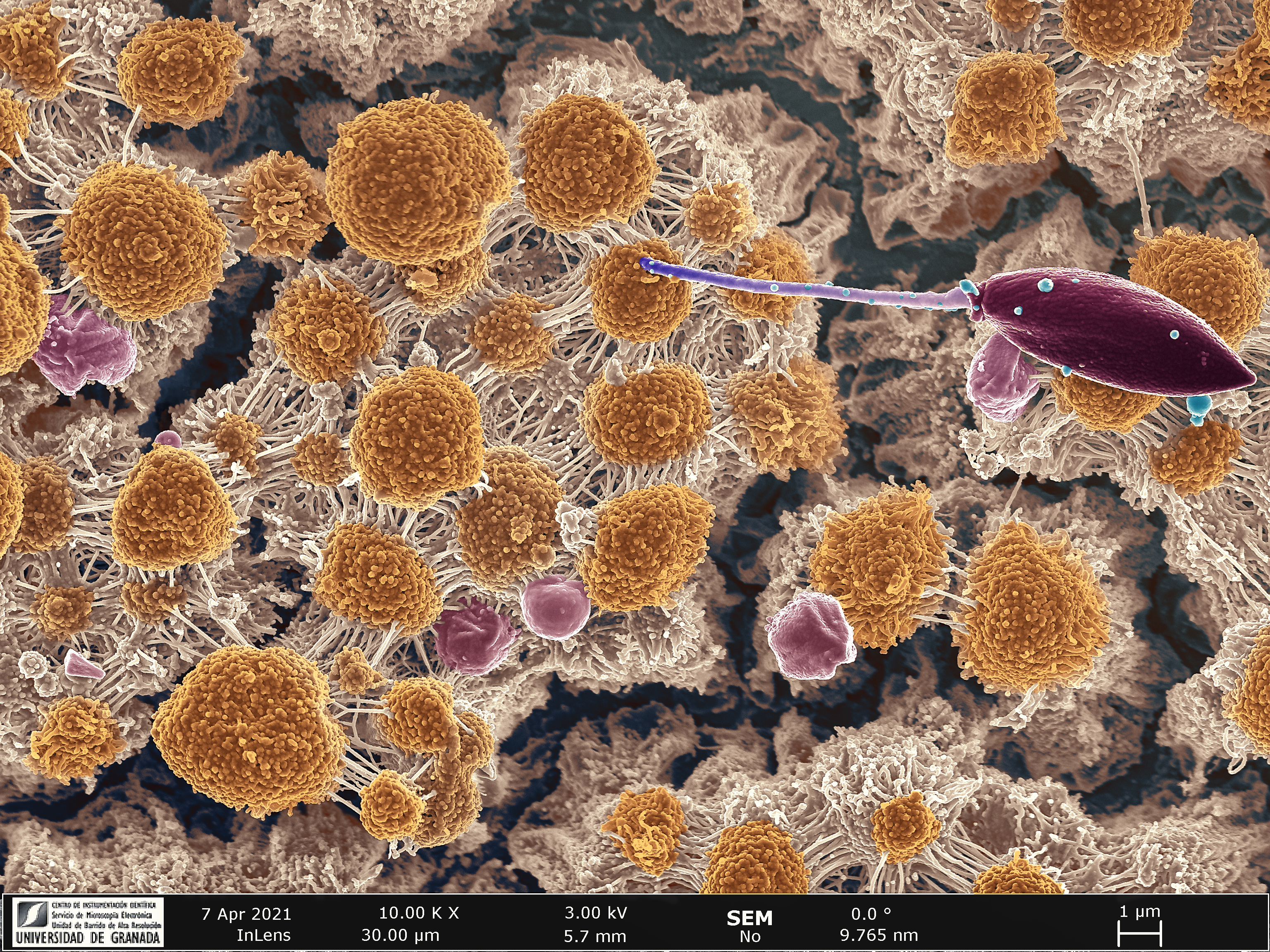
As commented above, these organisms can be relatively easily grown in liquid cultures. After observing their growth dynamics our team noticed the presence of a prolonged and stable stationary phase ( when nutrients are scarce) far longer than other human-infective trypanosomatid counterparts. More intrickingly, the giemsa stain of L. passim cultures at late logarithmic and stationary phase showed long and dense fibers of mucopolysaccharidic appearance and the presence of surface-attached cells lining at the air/liquid interface. Given these particular features, we analysed by different microscopy techniques the secreted polymeric material and also the surface attached cells. This charcterization led to the discovery of Extracellular Polymeric Substances (EPS) released by promastigote unicellular planktonic forms and the further developmental differentiation into a new parasitic lifestyle that we termed as trypanosomatid biofilms.
Biofilms are defined as surface-attached community of microorganisms embedded in a self-produced matrix of EPS composed mainly by exopolysaccharides but also nucleic acids (extracellular DNA and RNA), proteins, lipids and other biomolecules. This life cycle strategy is considered to be one of the most widely distributed and successful modes of life on the earth. However, the mechanisms of EPS secretion and biofilm formation have never been described in protozoan parasites.
After the structural analysis of L. passim EPS, we found patterns of repetitive monomeric units composed by exovesicles and polysaccharides that we termed as “spherulites''formed by long fibers and chains resembling “rosary beads''. This matrix was especially dense over surface-attached trypanosomatid biofilms in vitro, connecting individual cells and being a constitutive part of the architecture of such multicellular community. To ascertain whether or not trypanosomatid biofilms differentiated in vitro are also found in honeybees we developed a series of experimental infections with L. passim and found trypanosomatid biofilms in the hindgut of honeybees, demonstrating that developmental differentiation into this multicellular life cycle stages is a canonical developmental differentiation process in this parasite. Besides, one of the biochemical properties of trypanosomatid biofilms found in vitro was a higher resistance to Reactive Oxygen Species (ROS). Since the release of ROS to the gut lumen is one of the defense mechanisms against pathogens employed by insects, we think that this parasite innovative transformation could be implicated in the resilience of these parasites in bee hosts. Furthermore, we also found differentiation into biofilms in other honey bee trypanosomatids such as Crithidia mellificae, indicating that this parasite strategy could have evolved and maintained in more than one trypanosomatid species.

Finally, we also investigated whether or not such innovations in trypanosomatid parasites of honeybees could be advantageous upon osmotic pressures or cold-shock stress. As a result, we demonstrated that honeybee infective parasites are 2-3 fold more resistant than human-infective dixenous (two hosts) trypanosomatid parasites. The resistance to severe environmental changes inside and outside their hosts should be performed by evolving highly adaptable cells. In the case of trypanosomatid parasites, this is probably due to the protective role exerted by the release of EPS and the further ttansformation in biofilms.
These results set up the basis for the understanding of the successful expansion of this group of parasites in nature and to unveil the impact of such pathogens in honeybees, a keystone species in most terrestrial ecosystems.

Given that many questions remain unanswered such as what are the molecular mechanisms underlying developmental differentiation into trypanosomatid biofilms, what is the extension of such lifestyle in other trypanosomatids in nature or whether could this information could be used to hamper parasite transmission between honeybees, future research by our research team will be devoted to the molecular and cellular characterisation of this example of extreme biology.
References
- Buendía-Abad, M. et al. Workflow en of Lotmaria passim isolation: Experimental infection with a low-passage strain causes higher honeybee mortality rates than the PRA-403 reference strain. International Journal for Parasitology: Parasites and Wildlife 14, 68–74 (2021).
- Fantham, H. & Porter, A. Note on certain protozoa found in bees. J. Board. Agric. 19, (1911).
- Langridge, D. F. & McGHEE, R. B. Crithidia mellificae n. sp. an Acidophilic Trypanosomatid of the Honey Bee Apis mellifera. The Journal of Protozoology 14, 485–487 (1967).
Acknowledgments: We would like to acknowledge the Imaging Service of the Centro de Instrumentación Científica (Universidad de Granada) and specially Ms. Alicia González Segura and Dr. Lola Molina Fernández for picturing and post- processing of the Biofilm image referenced this article.
Follow the Topic
-
npj Biofilms and Microbiomes
The aim of this journal is to serve as a comprehensive platform to promote biofilms and microbiomes research across a wide spectrum of scientific disciplines.

Related Collections
With Collections, you can get published faster and increase your visibility.
Microbial endocrinology
Publishing Model: Open Access
Deadline: Oct 21, 2026
AMR and microbiome
Publishing Model: Open Access
Deadline: Apr 01, 2027



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in