Pathogenic bacteria hijack probiotic signal to activate virulence factor production and promote its survival
Published in Microbiology and Cell & Molecular Biology
Probiotic bacteria are live microorganisms that provide health benefits to the host upon consumption. Given the existing difficulties in treating human infections caused by antibiotic-resistant bacteria, the treatment or prevention of infectious diseases by the consumption of beneficial microbes has become an intriguing possibility. Numerous probiotics are already available for human consumption, and as a result, probiotics have become a multibillion-dollar industry. However, except for a few probiotics, the mechanistic basis for the claimed probiotic health benefits is sorely lacking, and more importantly, results from clinical trials testing the efficacy of probiotics remain unconvincing.
Infections caused by the human pathogen Streptococcus pyogenes, also known as GAS, remain a cause of significant human mortality and morbidity worldwide. GAS infections range from mild treatable throat infections (Pharyngitis or sore throat) to life-threatening invasive infections (sepsis and flesh-eating disease). Untreated recurring pharyngeal infections and the presence of GAS in human saliva are critical for disease transmission and the onset of more serious life-threatening invasive infections. Currently, there are no licensed human vaccines available, and as a result, GAS disease prevention is significantly hampered.
Streptococcus salivarius K12 (SAL) strain was originally isolated from the oral cavity of a healthy infant and sold as a commercial product (as “throat guard”) to prevent various oral diseases, including sore throat in school children caused by GAS infections. As for most commercial probiotic products, evidence for SAL health benefits is derived from in vitro GAS killing assays on agar plates. It was suggested that SAL uses two lantibiotics, salivaricin A and salivaricin B, to kill GAS and confer health benefits. However, critical in vivo evidence using either pre-clinical mouse models of GAS infection or human saliva ex vivo demonstrating their potency is sorely lacking. Furthermore, human clinical trials testing the efficacy of SAL probiotics against GAS infections offered conflicting results and remained inconclusive.
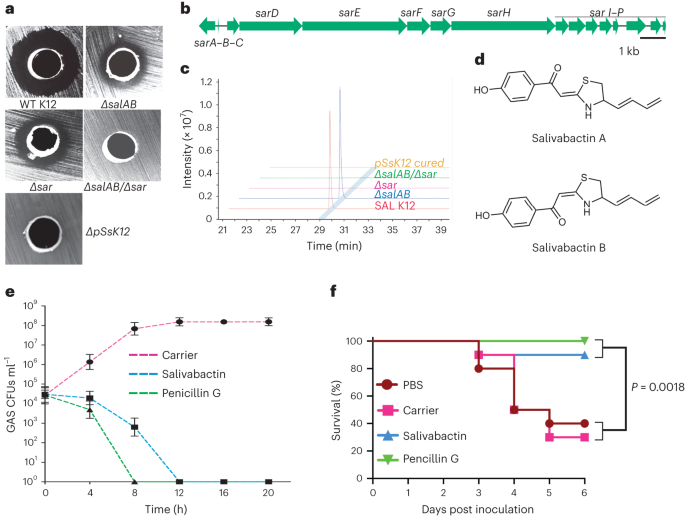
Discovery of a new antibiotic with a previously unknown chemical scaffold. Given the interest of our laboratory in devising new translational strategies to combat GAS infections, we tested the in vivo prophylactic efficacy of SAL against oropharyngeal GAS infections. As a first step, we have begun with testing the inhibitory role of salivaricins against GAS growth in vitro using salA/salB-inactivated double mutant strain. However, much to our surprise, the mutant strain still showed inhibition, albeit at lesser level. This result prompted us to consider the presence of a third SAL-derived previously unknown antimicrobial. In silico analyses of the SAL genome and megaplasmid sequences indicated the presence of a novel polyketide/non-ribosomal biosynthetic gene cluster (PK/NRPS-BGC) encoded in the megaplasmid that may be responsible for antimicrobial production. Consistent with this prediction, genetic inactivation of PK/NRPS-BGC abolished anti-GAS activity.
These findings clearly piqued our interest in the structure and function of the new antimicrobial, termed as salivabactin. However, the isolation and structure determination of small-molecule natural antimicrobials are beyond the expertise of our laboratory. Thus, we sought the expertise of Wenjun Zhang’s laboratory at the University of California, Berkeley, to elucidate the chemical identity of the compound. Using the mutant strains generated in our laboratory, Wenjun’s laboratory was able to purify salivabactin in milligram quantities from liters of SAL culture supernatant. Salivabactin is a relatively small molecule with a unique chemical scaffold. Using purified salivabactin, we demonstrated that it is potent against various gram-positive human pathogens but was relatively inactive against gram-negative pathogens. These findings also suggest the presence of gram-positive specific, potentially novel salivabactin target(s). More importantly, we were able to treat intramuscular invasive GAS infection in a mouse model by intramuscular administration of purified salivabactin.
Twist to the story. At this point, we were encouraged by the potential in vivo probiotic efficacy of SAL against GAS infections, largely due to the identification of a new potent SAL-derived antibiotic and the in vitro efficacy of SAL in inhibiting GAS growth. We proceeded to test the probiotic efficacy of SAL against GAS in an ex vivo model using human saliva and two in vivo mouse models mimicking GAS infection in the oropharynx and long-term mucosal colonization in the mouse vaginal lumen. However, contrary to our expectations, SAL was not only ineffective in preventing GAS infection but also promoted GAS persistence in human saliva and GAS colonization in vivo. Although we were surprised by the results, additional experiments confirmed the growth-promoting effects of probiotic SAL on GAS. Consequently, we set to sort out the potential mechanism by which SAL augments GAS survival ex vivo and in vivo.
To narrow down the possible mechanisms of GAS growth promotion by SAL, we first explored whether the phenotype was contact-independent or contact-dependent. SAL did not require direct contact with GAS for growth promotion as it promoted GAS growth even in the absence of direct contact in transwells. One possibility is that GAS produces protective proteases in the presence of SAL, which likely aids its evasion of antimicrobials and other hostile factors and promotes its survival. Because GAS produces several secreted proteases that contribute to its survival, we compared the expression levels of GAS proteases by qRT-PCR in the presence or absence of SAL. Of the five different secreted GAS proteases tested, we found drastic upregulation of an extracellular promiscuous protease SpeB only in the presence of SAL. SpeB is a major virulence factor that is known to degrade host-derived antimicrobials, immune factors and tissue matrix proteins, and promote disease pathogenesis. Intriguingly, the timing of speB upregulation during co-cultivation coincided with GAS growth promotion, suggesting a role for SpeB in increased GAS survival in the presence of SAL.
GAS eavesdrops SAL signal and trigger secreted protease production. Using saliva co-culture studies, we further confirmed that SAL induced early activation and robust induction of speB expression and secreted SpeB production. However, the key question regarding how GAS coordinates the early induction of SpeB production in the presence of SAL remained unanswered. A few years ago, our laboratory discovered that GAS coordinates speB expression in concert with high population density using a novel leaderless communication peptide (LCP) signal, SpeB-inducing peptide (SIP). Till this point, GAS SIP was the only documented LCP, and no other examples were available. Because a secreted peptide signal activates speB expression, we explored the possibility that a SAL-derived secreted signal may cross-activate speB expression in GAS during co-cultivation using cell-free culture supernatants from SAL growth. Consistent with our hypothesis, the secreted component of SAL growth induced speB expression, indicating the presence of a secreted SAL-derived activation factor. In GAS, LCP is produced, secreted, reinternalized, and recognized by the cytosolic receptor RopB, which activates speB expression via direct promoter interactions. We screened the SAL genome and megaplasmid sequences for RopB-like receptors and adjacently located LCP. Intriguingly, a ropB–LCP-like receptor–peptide pair was located immediately upstream of PK/NRPS-BGC and was involved in the regulation of salivabactin production. Using the tools developed during our original LCP discovery, we further demonstrated that secreted SAL-derived LCP is the basis for the cross-activation of speB expression in GAS. However, we found that the cross-activation is unidirectional as only SAL activated GAS gene expression, whereas GAS failed to influence SAL gene expression during co-cultivation. Interestingly, we found that both salivaricins are degraded by SpeB in vitro, which likely contributed to GAS growth promotion by preventing salivaricin-dependent GAS killing. Overall, we discovered a unique mechanism employed by the pathogen in which the pathogen eavesdrops a secreted signal produced by a probiotic to activate its virulence factor production. The proposed mechanism predicts that probiotic-derived secreted signal and pathogen-derived secreted toxin are the major players in interspecies interactions and augmentation of pathogen growth. Consistent with this, inactivation of either of these components abolished the promotion of pathogen growth by probiotic bacteria.
A hiccup along the way. Till this point, Hackwon Do and Nishanth Makthal spearheaded the project. Hackwon Do developed the tools and performed the major experiments. Both traveled to their respective home countries for different reasons, one to conduct faculty job interviews and the other to attend a family wedding. However, around the same time, the US imposed international travel bans due to the COVID-19 pandemic, and both could not travel back to the US for another 7 months. In the meantime, Hackwon accepted the job offer to start his own laboratory and stayed back in his home country. All these unexpected developments significantly delayed the completion of the final round of experiments. Almost a year later, Praveen Kumar Tripathi, a new postdoctoral fellow, joined the laboratory and restarted the project.
Devising a new translational strategy by cutting pathogen eavesdropping and augmenting probiotic antimicrobial production. Based on our findings, we concluded that SAL produces an LCP, which is used by GAS to produce secreted SpeB and promote its survival by degrading antimicrobials, including salivaricins. Thus, we proposed two genetic modifications in SAL to generate a more efficacious probiotic (engineered SAL, eSAL): i) genetic inactivation of SAL LCP to eliminate GAS eavesdropping and early SpeB production, and ii) coupling salivabactin production with a constitutive promoter to increase the production of SpeB-insensitive salivabactin. Collectively, we reasoned that delayed SpeB production, reduced degradation of salivaricins, and increased salivabactin production will result in the inhibition of GAS growth by SAL. In vitro and in vivo characterization of eSAL confirmed that genetic modifications in eSAL resulted in the following expected outcomes: delayed and reduced SpeB production, increased salivabactin production, increased GAS killing, and reduced GAS survival in mouse models of infection.
Summary. In summary, results from work conducted in 3 different laboratories, Kumaraswami, Zhang, and La Rock’s laboratories, highlighted the significance of studying probiotic efficacy in human-related environments and understanding the mechanistic basis of interspecies interactions between probiotic and pathogenic bacteria. Elucidation of the molecular details of interspecies interactions promoted the discovery of a novel antibiotic as well as a redesigned, more efficacious probiotic to treat human bacterial infections in preclinical mouse models. We hope that our findings will prompt detailed investigations into the in vivo efficacy of other probiotic bacteria, which may yield improved tools to combat drug-resistant bacterial infections.
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in