PrPC controls epithelial-to-mesenchymal transition in EGFR-mutated NSCLC: implications for TKI resistance and patient follow-up
Published in Cancer

Where did we start?
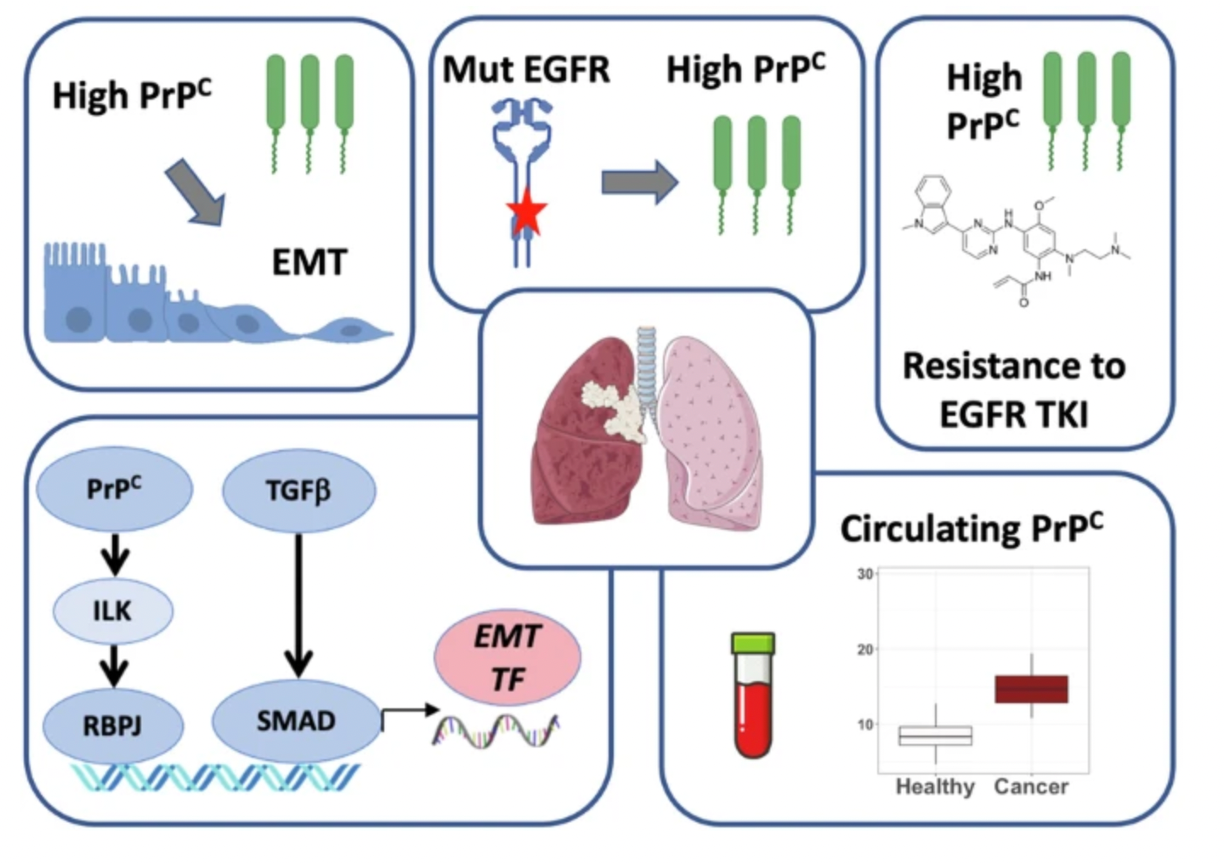
Lung cancer remains the deadliest cancer globally, accounting for 1.8 million deaths in 2020, which corresponds to 18% of all cancer-related deaths1. The most common druggable genetic alteration found in Non-Small Cell Lung Carcinomas (NSCLC) are EGFR mutations, occurring in 15–25% of cases among Caucasians, and up to 40–55% in East Asians2. Patients with EGFR-mutated (EGFRm) NSCLC benefit from treatment with tyrosine kinase inhibitors (TKI) targeting EGFR. However, despite advancements in patient care, particularly with the third-generation TKI osimertinib, all patients treated with these TKI develop resistance after 9 to 12 months of treatment3,4. Resistance mechanisms to TKI include several processes, one of which is Epithelial to Mesenchymal Transition (EMT)5,6. During EMT, epithelial cells acquire mesenchymal characteristics, adopting a wide spectrum of phenotypes depending on the extent of mesenchymal features gained compared to the epithelial traits retained7. While EMT is a physiological process, it is also a well-recognized mechanism in cancer cells, especially in the context of resistance to anticancer treatments7. Unfortunately, the signals and effectors orchestrating EMT in NSCLC remain poorly understood8. Recent work from our laboratory has shown that the expression of the cellular prion protein (PrPC)-encoding gene PRNP is specifically enriched in the mesenchymal subtype of colon cancer and regulates the expression of the master EMT transcription factor (TF) ZEB19. The PrPC protein is known for its role in spongiform encephalopathies, such as Creutzfeldt-Jakob disease, which occurs when PrPC adopts a pathological conformation known as the Scrapie form: PrPSc10. In line with our research, PrPC has emerged over recent years as a significant contributor to EMT in various other cancer types and has been implicated in several cancer hallmarks11. Moreover, multiple studies have reported the pro-migratory and pro-invasive properties of PrPC in cancer cells11. Notably, in lung cancer, Lin et al. have demonstrated that PrPC is more abundantly expressed in invasive versus non-invasive cell lines and enhances lamellipodium formation12. Despite these findings, our understanding of the interplay between PrPC, EMT, and resistance to anti-EGFR TKI in lung cancer remains incomplete, which is why we have undertaken this research (Figure 1).

Figure 1
What did we do?
As a first step, we conducted a comprehensive analysis by mining several public datasets of lung adenocarcinoma (LUAD) cell lines and patient samples to identify molecular signatures associated with PRNP expression and EMT in LUAD. We then utilized cell-based assays, particularly loss-of-function approaches, on three EGFRm LUAD cell lines to investigate the role of PRNP in regulating EMT and to explore the interplay between PrPC and EGFR. Finally, we integrated in silico analyses, in vitro assays, and plasma measurements of circulating PrPC in patients undergoing treatment for LUAD to assess the contribution of PrPC to TKI resistance in EGFRm LUAD (Figure 2).

Figure 2
What did we find?
The analysis of five datasets from LUAD cells and patients demonstrated a strong correlation between PrPCand EMT in NSCLC. PRNP expression was consistently associated with EMT across all five datasets and was linked to several EMT-TF. Furthermore, we observed a robust positive correlation between PRNPexpression and mesenchymal scores, which reflect the EMT process, as defined by other researchers. In the Onco-HEGP cohort, PRNP mRNA levels were significantly higher in mixed or mesenchymal tumors compared to epithelial tumors. Subsequently, we established a functional relationship between PrPC and EMT through cell-based experiments. Specifically, using loss-of-function approaches, we found that PrPC is essential for maintaining mesenchymal features and is required for cells to acquire mesenchymal characteristics in response to growth factors such as TGFβ. We further demonstrated that PrPC exerts its effects via the NOTCH pathway, specifically through an ILK-RBPJ axis which works in conjunction with TGFβ signaling. Both in silico and in vitro approaches revealed a functional and reciprocal interaction between EGFR and PrPC. Our in silico analysis indicated an association between PRNP expression and the enrichment of a signature linked to EGFR activation. Additionally, analysis of LUAD cell line datasets showed higher PrPC expression in EGFRm cells compared to EGFR wild-type (WT) cells. This observation was corroborated by our analysis of multiple patient datasets, where PRNP levels were elevated in EGFRm tumors at both transcriptomic and proteomic levels. At the cellular level, exposure to EGF increased PrPCexpression at both mRNA and protein levels. We also identified a reciprocal relationship between PrPC and EGFR expression, where each influenced the other, and PrPC appeared to regulate EGFR activation in LUAD. Given that PrPC has been implicated in resistance to anticancer therapies across various cancer types11, we investigated whether PrPC, which promotes EMT—a known mechanism of resistance to anti-EGFR therapies—could contribute to resistance to anti-EGFR treatments in NSCLC. Notably, higher levels of PrPC were associated with resistance to anti-EGFR TKI in several published cell datasets. Moreover, our cellular experiments demonstrated that osimertinib was more effective when PrPC was silenced. To explore the clinical implications of these findings, we measured PrPC levels in plasma samples from two cohorts of NSCLC patients treated at the European Hospital Georges Pompidou. We discovered elevated PrPC levels in the plasma of EGFRm NSCLC patients and observed that longitudinal analysis of PrPC plasma levels during different stages of clinical management in several patients could reflect disease progression (Figure 3).

Figure 3
What is next?
Our research has indicated a potential role for PrPC in the resistance to anti-EGFR TKI in EGFR-mutated NSCLC, that needs to be validated in larger series. Considering that PrPC can be targeted, we could explore whether inhibitors could effectively restore sensitivity to anti-EGFR TKI in EGFRm NSCLC, particularly in vivo. Furthermore, plasma PrPC could be a surrogate marker of progression in patients with EGFRm lung cancer and should be further evaluated.
References
1 Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2021; 71: 209–249.
2 Yang C-Y, Yang JC-H, Yang P-C. Precision Management of Advanced Non-Small Cell Lung Cancer. Annu Rev Med 2020; 71: 117–136.
3 Rotow J, Bivona TG. Understanding and targeting resistance mechanisms in NSCLC. Nat Rev Cancer 2017; 17: 637–658.
4 Passaro A, Jänne PA, Mok T, Peters S. Overcoming therapy resistance in EGFR-mutant lung cancer. Nat Cancer 2021; 2: 377–391.
5 Bronte G, Bravaccini S, Bronte E, Burgio MA, Rolfo C, Delmonte A et al. Epithelial-to-mesenchymal transition in the context of epidermal growth factor receptor inhibition in non-small-cell lung cancer. Biol Rev Camb Philos Soc 2018; 93: 1735–1746.
6 Zhu X, Chen L, Liu L, Niu X. EMT-Mediated Acquired EGFR-TKI Resistance in NSCLC: Mechanisms and Strategies. Front Oncol 2019; 9: 1044.
7 Weng C-H, Chen L-Y, Lin Y-C, Shih J-Y, Lin Y-C, Tseng R-Y et al. Epithelial-mesenchymal transition (EMT) beyond EGFR mutations per se is a common mechanism for acquired resistance to EGFR TKI. Oncogene 2019; 38: 455–468.
8 Tulchinsky E, Demidov O, Kriajevska M, Barlev NA, Imyanitov E. EMT: A mechanism for escape from EGFR-targeted therapy in lung cancer. Biochim Biophys Acta Rev Cancer 2019; 1871: 29–39.
9 Le Corre D, Ghazi A, Balogoun R, Pilati C, Aparicio T, Martin-Lannerée S et al. The cellular prion protein controls the mesenchymal-like molecular subtype and predicts disease outcome in colorectal cancer. EBioMedicine 2019; 46: 94–104.
10 Oesch B, Westaway D, Wälchli M, McKinley MP, Kent SB, Aebersold R et al. A cellular gene encodes scrapie PrP 27-30 protein. Cell 1985; 40: 735–746.
11 Mouillet-Richard S, Ghazi A, Laurent-Puig P. The Cellular Prion Protein and the Hallmarks of Cancer. Cancers 2021; 13: 5032.
12 Lin S-C, Lin C-H, Shih N-C, Liu H-L, Wang W-C, Lin K-Y et al. Cellular prion protein transcriptionally regulated by NFIL3 enhances lung cancer cell lamellipodium formation and migration through JNK signaling. Oncogene 2020; 39: 385–398.
Follow the Topic
-
Oncogene
This journal aims to make substantial advances in our knowledge of processes that contribute to cancer by publishing outstanding research.

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in