Selecting the most appropriate SpCas9 variants for your own interests
Published in Bioengineering & Biotechnology
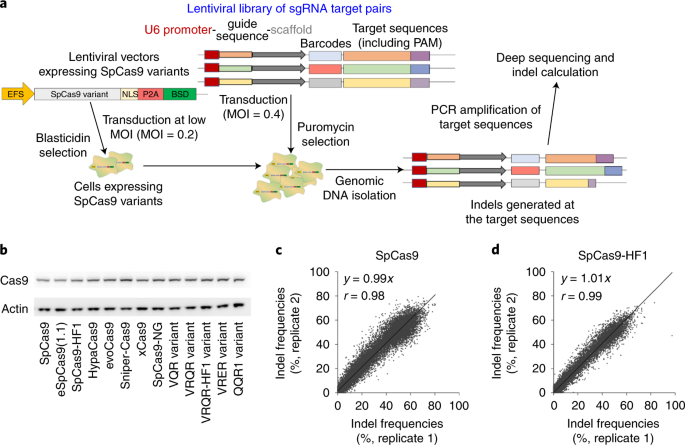
CRISPR-Cas9 system, especially Streptococcus pyogenes Cas9 (SpCas9), has been widely used in various fields due to its effectiveness and convenience1. However, there are limitations for broader applications as SpCas9 systems induce not only high on-target activities but also exhibit relatively high off-target activities. They also require specific PAM sequences for target recognition that limit its surveillance of the genome. Several groups have reported variants of the SpCas9 to improve its high-fidelity or PAM variants for the purpose of minimizing its potentially harmful off-target activity or expanding its repertoire of targetable PAM sequences, respectively. However, there are too many choices; we do not know which SpCas9 variant we should use for a genome editing at a given target sequence.
In order to address these limitations, we comprehensively compared 13 SpCas9 variants including the wild-type SpCas9 (listed below).
- PAM variants : Wild-type SpCas9, VQR variant4, VRER variant4, VRQR variant5, VRQR-HF1 variant5, QQR1 variant6, xCas97 and SpCas9-NG8
- High-fidelity variants : Wild-type SpCas9, eSpCas9(1.1)9, SpCas9-HF15, HypaCas910, evoCas911, xCas9 and Sniper-Cas912
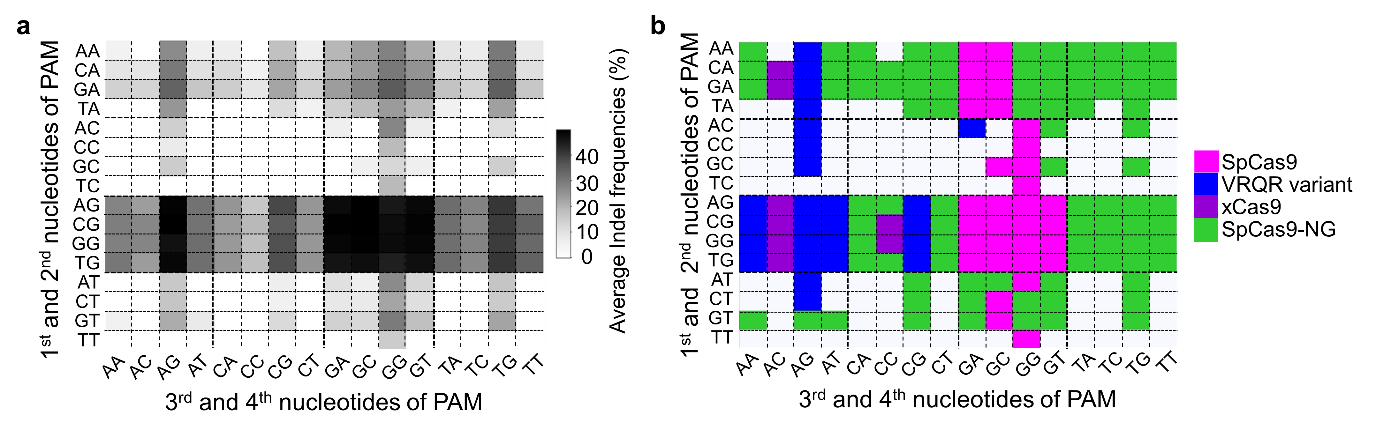
Initially, all possible 4-nt PAM sequences with 30 protospacers were evaluated for PAM compatibilities. After defining indel frequencies that were greater than 5% as functional PAM sequences, we were able to determine that 156 PAM sequences (156 = 89 by SpCas9-NG + 33 by SpCas9 + 26 by VRQR variant + 8 by xCas9) could be targeted with 4 distinct PAM variants from 8 variants that we examined (Fig. 1).
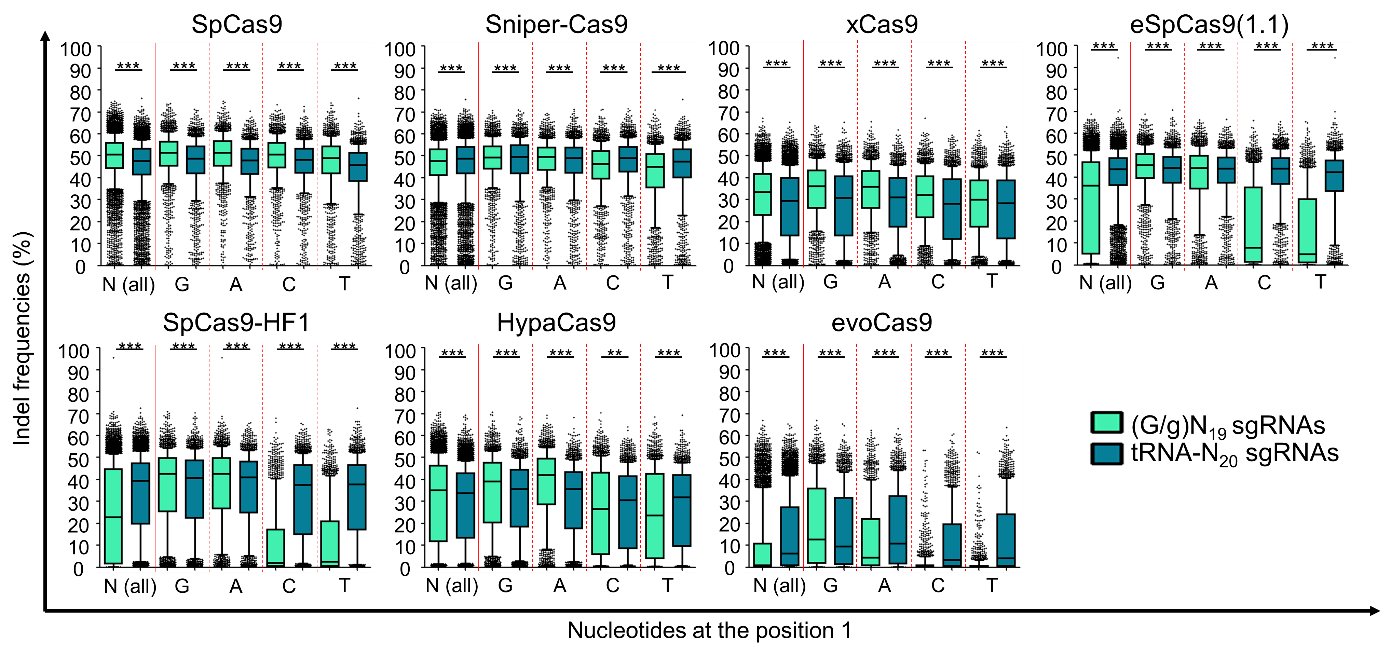
Next, we utilized two sgRNA expression approaches by a U6 promoter to compare on-target activities of high-fidelity variants: (G/g)N19 sgRNAs, 20-nt guide sequences with a G at the 5' terminus that we matched (GN19) or mismatched (gN19) to the target sequences, and tRNA-processed N20 guide sequences (tRNA-N20 sgRNAs) that were perfectly matched to the target sequences. Using high-throughput analysis, we demonstrated that a proper guide RNA expressing system should be used to establish high on-target activities depending on the variants and first nucleotide of the target sequences (Fig. 2).
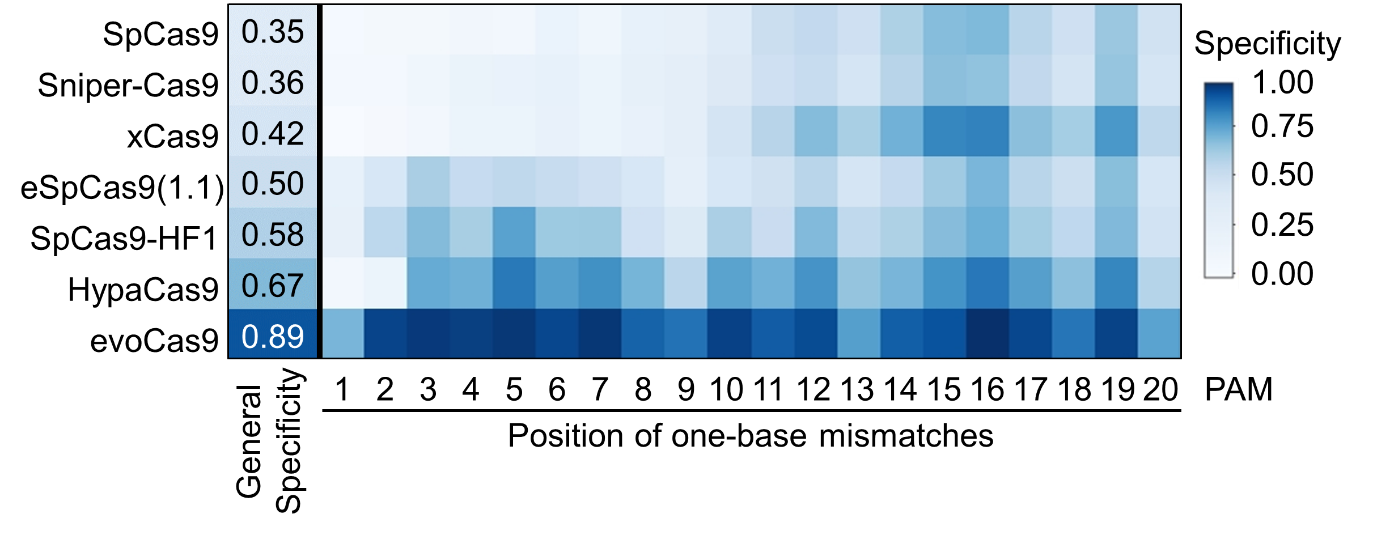
Using 1-bp mismatched target sequences, we compared the specificity of each high-fidelity variant, and found that the highest and lowest specificities were showed by evoCas9 and SpCas9, respectively. Interestingly, unlike with SpCas9, eSpCas9(1.1), SpCas9-HF1, HypaCas9 and evoCas9 exhibited two major peaks of intolerance to mismatches in both PAM-proximal and distal regions (Fig. 3). Based on our results, we developed a computational tool that predicts the activities of SpCas9 variants.
Overall, the systematic and comprehensive comparison of all variants simultaneously provided many challenges. Conducting all experiments in controlled conditions for each variant was very difficult and required immense focus, patience and endurance. Nonetheless, we were confident that our comprehensive results would assist future researchers with their research goals. We hope that our findings and the prediction tool may provide useful insights for future works and help expand our understanding of genome editing.
Thank you for Jinman Park and Hyongbum Henry Kim for helping me with this blog.
Our paper can be found with the following link: https://www.nature.com/articles/s41587-020-0537-9
- Hsu, P.D., Lander, E.S. & Zhang, F. Development and applications of CRISPR-Cas9 for genome engineering. Cell 157, 1262-1278 (2014).
- Kim, H.K. et al. High-throughput analysis of the activities of xCas9, SpCas9-NG and SpCas9 at matched and mismatched target sequences in human cells. Nature Biomedical Engineering (2020).
- Wang, D. et al. Optimized CRISPR guide RNA design for two high-fidelity Cas9 variants by deep learning. Nature communications 10, 4284 (2019).
- Kleinstiver, B.P. et al. Engineered CRISPR-Cas9 nucleases with altered PAM specificities. Nature 523, 481-485 (2015).
- Kleinstiver, B.P. et al. High-fidelity CRISPR-Cas9 nucleases with no detectable genome-wide off-target effects. Nature 529, 490-495 (2016).
- Anders, C., Bargsten, K. & Jinek, M. Structural Plasticity of PAM Recognition by Engineered Variants of the RNA-Guided Endonuclease Cas9. Molecular cell 61, 895-902 (2016).
- Hu, J.H. et al. Evolved Cas9 variants with broad PAM compatibility and high DNA specificity. Nature 556, 57-63 (2018).
- Nishimasu, H. et al. Engineered CRISPR-Cas9 nuclease with expanded targeting space. Science 361, 1259-1262 (2018).
- Slaymaker, I.M. et al. Rationally engineered Cas9 nucleases with improved specificity. Science 351, 84-88 (2016).
- Chen, J.S. et al. Enhanced proofreading governs CRISPR-Cas9 targeting accuracy. Nature 550, 407-410 (2017).
- Casini, A. et al. A highly specific SpCas9 variant is identified by in vivo screening in yeast. Nature biotechnology 36, 265-271 (2018).
- Lee, J.K. et al. Directed evolution of CRISPR-Cas9 to increase its specificity. Nature communications 9, 3048 (2018).
Follow the Topic
-
Nature Biotechnology
A monthly journal covering the science and business of biotechnology, with new concepts in technology/methodology of relevance to the biological, biomedical, agricultural and environmental sciences.



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in