Small body size is associated with increased evolutionary lability of wing skeleton proportions in birds
Published in Ecology & Evolution and Zoology & Veterinary Science
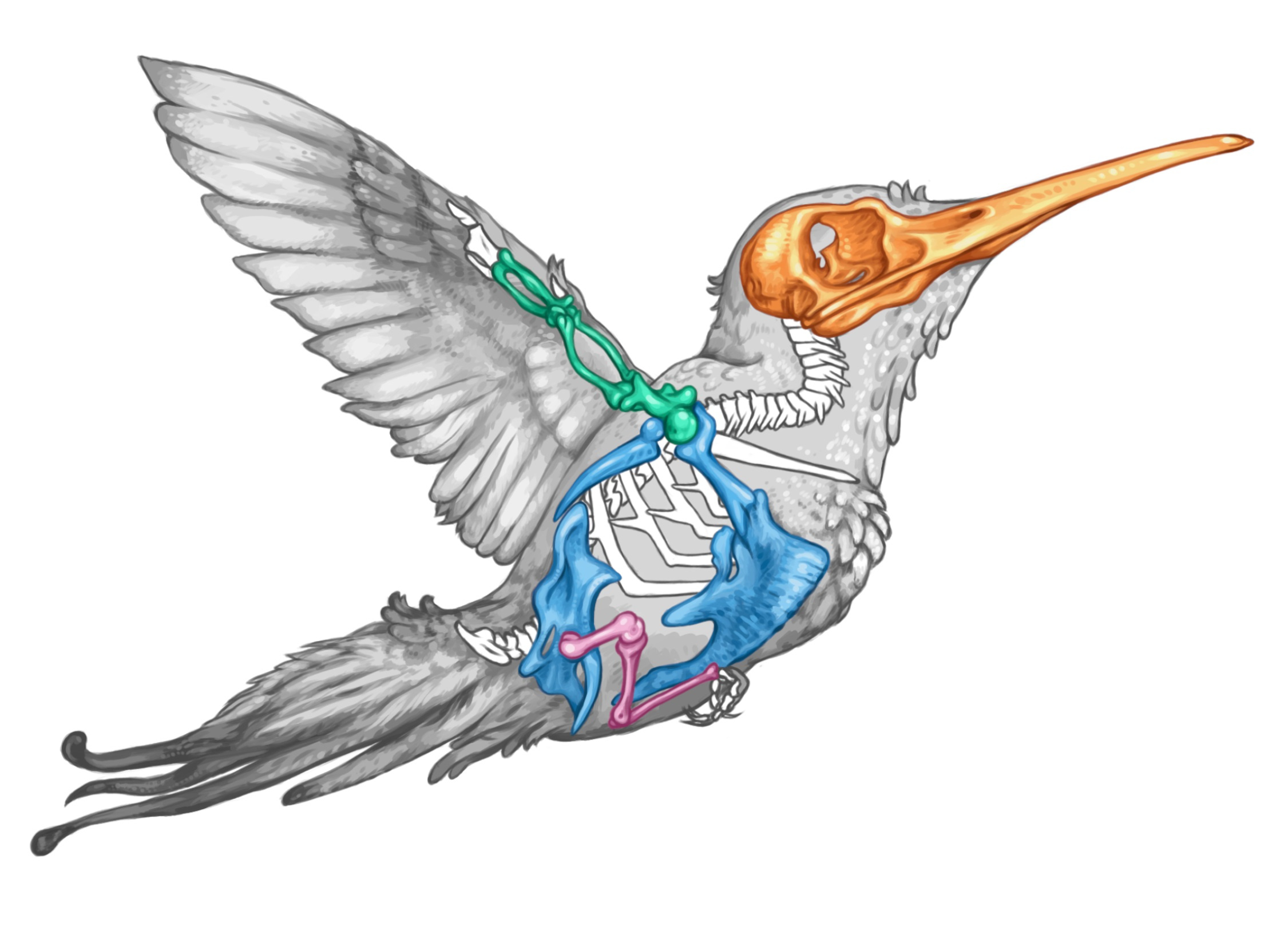
"Modular" evolution:
Birds have 'modular' skeletons, in which different body regions- such as wings, legs and head, can evolve semi-independently. The investigation of evolutionary modules and their influence on evolutionary change is a key avenue of research in biology.
Patterns of organised evolution of different traits across organism body plans, which may be imposed by development, mechanical or functional demands, are thought to be both a reflection and key driver of long term evolutionary change. Whether traits become allied and evolve together as a single 'module', or diverge independently, may determine the fate of entire lineages. However, disentangling developmental, mechanical and functional causes of 'modular' evolution remains a major challenge in evolutionary biology.
Flying birds' wings: a model system
Wings have a similar developmental history and function across flying birds, but larger birds' wings are under greater mechanical stress than small birds because of their larger body masses.
Birds vary in body mass by a factor of 10,000, and the modular evolutionary layout of their skeletons is already well-described[1,2]. Birds are therefore a fantastic system with which to unravel the contributions of different possible drivers of modular evolution. Previous researchers have suggested that traits within locomotory structures placed under great strains should be expected to evolve together due to mechanical demands for structural reinforcement.
We therefore hypothesised that the relative sizes of individual bones within the avian wing should evolve less independently in bird lineages with large bodies, and should be freer to evolve more independently of one another in smaller birds.
Approach:
We downloaded a large open-access dataset of 228 bird species’ skeletal proportions (Navalón et al., 2022[3]), representing a diversity of bird lineages, different ecological niches and ranging in body mass from a dainty 3 grams (Archilocus colubris, the Ruby-throated hummingbird) to a monstrous 11 kilograms (Vultur gryphus, the Andean condor). We tested whether different pairs of bones tended to evolve together by mapping changes in bone proportions across a family tree stitched together from publicly available phylogenies that represent the history of evolutionary relationships between birds. We isolated the influence of body mass upon the evolutionary alliance of different traits within evolutionary modules in two ways:
1) We arranged all of Navalón’s birds from smallest to largest, and divided the data into many different cohorts, comparing the patterns of modular evolution we found between them.
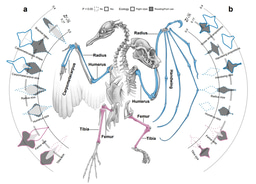
2) We compared how strongly different pairs of traits in the wing, head and leg evolved together, across the entire dataset. Then we asked whether the differences between predicted trait co-evolution and observations tended to be larger or smaller in birds with different body masses. The patterns we expected to find are illustrated in Figure 1.

Left: We expect Method 1 to reveal stronger co-evolution between wing bones in larger birds. Right: We expect Method 2 to demonstrate that smaller taxa (points) are scattered more widely from overall trends of evolutionary associations between traits (Dm) in the wing. By contrast, there should be no pattern in the head. If these patterns are significant, we expect observations (solid oval of right-hand ovals) to show a characteristically different distribution to a simulated ‘null’ model of evolution (filled oval).
Results:
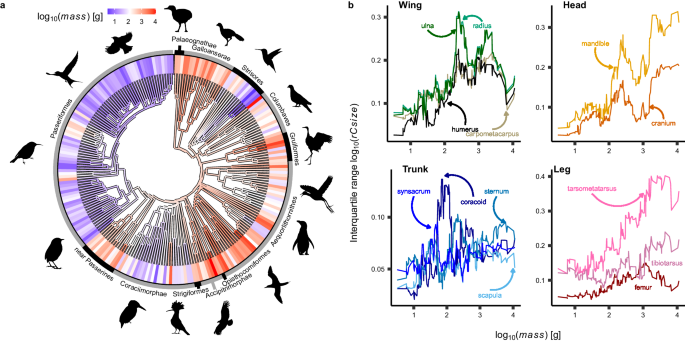
Our results followed our predictions; both methods converged to show that the allied evolution of sizes of bones within the head and leg is unaffected by variation in body mass, confirming that our methods do not produce unexpected ‘false positives’. By contrast, we found strong support demonstrating that the relative sizes of bones within the wing evolve together more closely in large birds- just as expected (Figure 2). This means that individual wing bones are able to evolve more freely of one another in smaller birds. We suggest that this may explain why novel styles of flight- like the dynamic soaring of hyper-aerial swifts, bounding flaps of perching birds, and hovering or hummingbirds and sunbirds, tend to emerge in small-bodied bird lineages. Our work provides a blueprint demonstrating that it is possible to untangle the contributions of functional, mechanical and developmental controls on the establishment of and changes to evolutionary modules, allowing us to deepen our understanding of the evolutionary story of birds- and perhaps many other groups.

Left: Method 1 shows that traits within the wing become more allied in larger birds, that there is no substantial change in the head or leg, and that trait evolution becomes more independent in the trunk (body) in larger birds.
Right: Method 2 Shows that individual species with small bodies are more likely to evolve extreme and novel combinations of wing traits (top), and the observed distribution (solid contour) is characteristically different from the null (filled), evidence that this trend is significant. By contrast, the strength of trait integration in the head (bottom) does not vary between small and large birds. The observed and null distributions overlap. Point colour distinguishes major bird groups.
Future directions:
Postdoctoral Researcher Dr. Andrew Orkney reflects on the importance of open access publishing and data sharing. “Our study was made possible by support from other scientists, who made available datasets of bird skeletal properties and evolutionary relationships. This work demonstrates the importance of sharing and actively disseminating datasets, engaging and supporting other scientists– especially early career researchers,”
Prof. Brandon P. Hedrick is keen to explore how similar evolutionary patterns may, or may not, exist in bats.
"Other than birds, bats are the only modern group to have evolved powered flight. Understanding whether there is one pathway or multiple different pathways to flight will greatly increase our understanding of this fantastic innovation for traversing environments."
Postdoctoral Researcher Dr. Priscila Rothier, who recently joined the lab, wants to further disentangle the web of interactions between development, adaptation and mechanical limitation, that contribute to modularity:
“Birds vary not only in body mass, but also in their developmental strategies. While some species, like passerines (garden birds), are highly altricial and heavily rely on parental care, others, such as ducklings, display early precocial activity. Because developmental processes are known to impact trait integration, one would expect that the diversity in hatchling development would also influence how skeletal elements covary and evolve.”
[1] Orkney et al., 2021: https://doi.org/10.1038/s41559-021-01509-w
[2] https://communities.springernature.com/posts/patterns-of-skeletal-integration-in-birds
[3] Navalón et al., 2022: https://doi.org/10.1038/s41586-022-05372-y
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in