Some dinosaurs liked it hot!
Published in Ecology & Evolution

Where are all the polar long-necked, giant sauropod dinosaurs? Dinosaurs dominated Mesozoic terrestrial ecosystems globally for almost 160 million years (from ~230–66 million years ago). They radiated into a wide range of ecotypes, dietary habits, and body sizes, the latter including those for the largest land animals of all time. Although dinosaurs show a cosmopolitan distribution, several groups seem to be restricted to different latitudinal bands. Whereas the mostly carnivorous dinosaurs (think of T. rex, Velociraptor and kin) and herbivorous ornithischians (like Triceratops, Stegosaurus, and the many duck-billed dinosaurs) seemed almost ubiquitously present, with many representatives recovered in polar Alaska and Antarctica, the long-necked behemoths of the Mesozoic, sauropods, seemed to prefer lower, more tropical latitudes. The question for palaeontologists was then: why did a pole-to-pole geographic distribution characterize ornithischians and theropods, while sauropods were restricted to lower latitudes?

The reconstruction of a Late Cretaceous, high latitude dinosaur bearing locality (Prince Creek Formation in the North Slope of Alaska) portrays the dinosaur more common in these sites, showing ornithischians (the ceratopsian Pachyrhinosaurus is in this figure) and theropods (the dromaeosaurids depicted here).
But where are the sauropods? Figure from Chiarenza et al. 2020.
We know that the geographic distributions of extant species largely reflect their environmental tolerance. For example, today birds and mammals have a near-global distribution, extending into high, polar latitudes, whereas the diversity of most reptiles, such as lizards, snakes, turtles, and crocodylians, as well as amphibians, are primarily concentrated in lower latitudes, closer to the Equator. This question partially overlaps with another, wider enigma concerning dinosaur biology, which has been debated ever since the recognition in 1842 of dinosaurs as a unique group of organisms.

Cover of Adrian Desmond's "The hot-blooded dinosaurs: a revolution in palaeontology", the influential scipop book vulgarising the movement triggered by palaeontologists John Ostrom and Robert Bakker in the 1960's re-evaluating dinosaurs as active, possibly warm-blooded animals. The older, opposite view that was considered as the standard model for most of the early XX century, treated dinosaurs as ectothermic-bradymetabolic animals closer to the classic reptilian stereotype. Modern biologists are of course now aware that vertebrate physiology is not as black and white as the narrative of "scientific revolutions" oftentimes portrays...
During the Ostrom/Bakker-led revolution of the ‘60s, dinosaurs were re-conceived as active, ‘sophisticated’ animals. This view was opposed to the stereotypical lumbering, reptilian attitude that was imposed on these animals and was used to portray them for most of the first half of the XX century. Although many of these attributions are quite subjective and unsubstantiated in modern science, dinosaur physiology, whether they were "cold" or "warm blooded", has been a fundamental question on dinosaur biology attracting both researchers and media attention. Since the ubiquitous geographic distribution of birds and mammals partially reflects their endothermic, active physiology, while cold-blooded reptiles and amphibians are largely restricted to the tropics, with every new latitudinally extreme discovery of non-avian dinosaur sites, re-evaluations and implications for the physiology of the group were made. Philip Mannion, an expert on sauropod dinosaurs and one of my supervisors during my PhD, noticed an interesting pattern regarding the distribution of his favourite critters: during the Late Cretaceous (100–66 Ma) there is evidence for distinctive patterns among the three main dinosaurian subclades, with sauropods more latitudinally restricted than ornithischians and theropods.
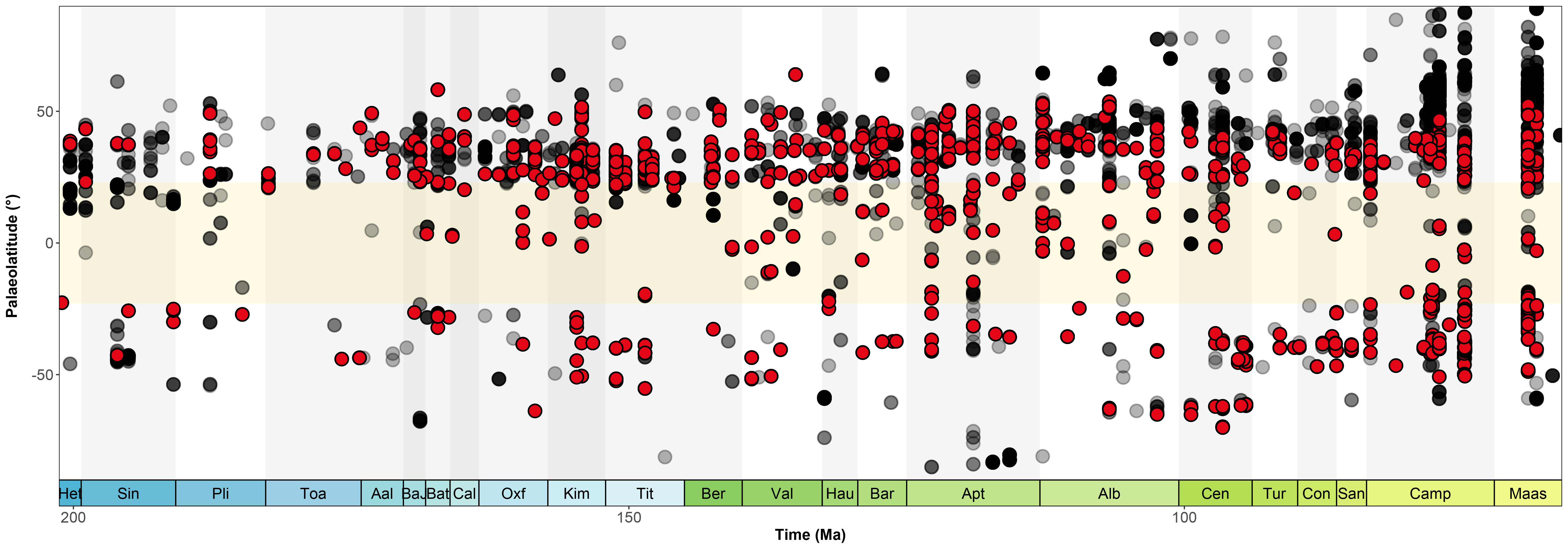
Palaeolatitudinal distribution of dinosaur occurrences through the Jurassic-Cretaceous interval: the black dots represent non-sauropod dinosaur occurrences while the red ones are sauropods. You can easily notice how non-sauropod dinos stretch out further North and South than do sauropods since the Early Jurassic up to their extinction 66 million years ago.
In particular, sauropod dinosaurs apparently peaked in diversity at southern palaeolatitudes of ~40–50º, with their remains entirely unknown from palaeolatitudes greater than ~66° in either hemisphere. By contrast, the apparent peak in ornithischian diversity corresponds to higher palaeolatitudes (~50–70º) in both hemispheres. If correct, this might indicate broad biological differences between sauropods and other dinosaurs. That study analysed the distribution and preservational patterns of the dinosaurian fossil record, but it did not examine dinosaur distributions in the context of palaeoclimate, and therefore could not address underlying questions regarding the potential implications of understanding dinosaur physiological diversity.

Sauropods were impressive animals and clearly par excellence dinosaurs. Here Alamosaurus, one of the last of the sauropods, dwarves his contemporary T. rex. The author of this post is highlighted by the red arrow and serves as a human scale.
In a new study published today in the journal Current Biology, we analysed the distribution of dinosaurs through the Mesozoic, applying several methods to evaluate whether the occurrence of fossil findings truly reflects the distribution of dinosaurs, after accounting for the vagaries of the fossil record, and whether some specific environmental drivers, such as climate, might have affected their distributional patterns. Because of the risk of reading the dinosaurian fossil record too literally and reconstructing dinosaur diversity and distribution from partial data, some authors have suggested that the spatial patterns are indeed artefactual. In this new study, my co-authors and I employed coverage-based subsampling approaches to evaluate the degree to which equivalently sampled palaeo-latitudinal areas are comparable, before using Earth System and ecological modelling techniques to directly quantify how environmental drivers (e.g. past temperature and precipitation) might have affected dinosaur distribution. It was vital for this project to have the collaboration of Alex Farnsworth, a leading palaeoclimatologist who has already worked with me on two publications using climate and ecological modelling to investigate palaeobiological questions concerning the dinosaurs. Alex provided his palaeoclimatological knowledge and the palaeoclimate data from the Bridge group in Bristol. We also collaborated with Matthew Carrano, an expert on theropods and Mesozoic dinosaur diversity more broadly, and who is the main contributor for dinosaur data in the open source dataset “The Paleobiology Database”, which comprises an essentially comprehensive record of the published dinosaur literature. Matt, Phil, and I checked, updated, and revised a large proportion of these data prior to analysis. Finally, whilst working on this project I was hired as a postdoc by Sara Varela, a palaeoecologist who is also one of the leading expert on species distribution modelling and the application of such macroecological techniques in deep time. This resulted in the ideal combination of people and skills to finally test the hypothesis of how palaeoclimate might have interacted with dinosaur biogeography and physiological diversity.
Our study reveals that sauropod dinosaurs were more strongly affected by temperature-related barriers during their evolutionary history than other dinosaurs, i.e. theropods and ornithischians. This phenomenon was particularly marked in the cold seasons, with temperature ranges for sauropods skewed towards higher temperatures than in the other dinosaurs. The richest sauropod localities seem to have coincided with habitats characterised by warmer, semi-arid environments, whereas theropods and ornithischians could endure even the harshest winters of the North Pole. What was the reason for this more pronounced affinity for warmer temperature in sauropods compared to other dinosaurs? Maybe high-latitude, colder regions were thermally hostile to the maintenance of viable sauropod populations, and these dinosaurs were simply unable to thrive in such settings? Perhaps sauropods were physiologically capable of living in high latitude environments, but were just much poorer at doing so than their dinosaurian cousins? The intriguing pattern of higher sauropod diversity at lower palaeolatitudes, and their absence from high latitude dinosaur assemblages, particularly during cooler intervals and in the Northern Hemisphere, raises the question of whether sauropod physiology was most efficient in warmer environments.
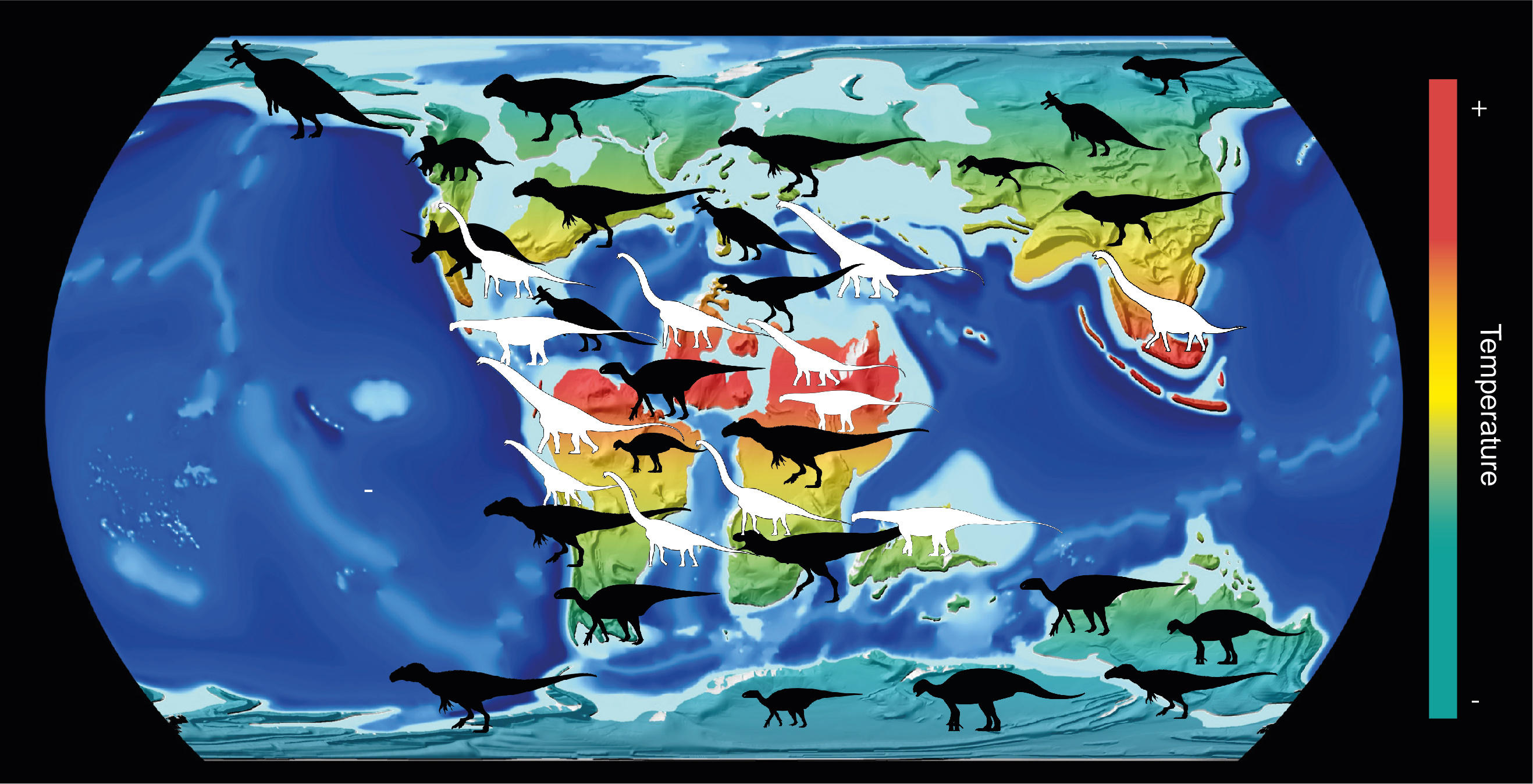 Graphical abstract for our paper, where the more latitudinally restricted distribution of sauropod dinosaurs (white silhouettes) is compared to the wider, pole-to-pole distribution of ornithischians and theropods.
Graphical abstract for our paper, where the more latitudinally restricted distribution of sauropod dinosaurs (white silhouettes) is compared to the wider, pole-to-pole distribution of ornithischians and theropods.
Remember the first scene depicting a glorious CGI dinosaur in Jurassic Park? The palaeontologist, Dr Alan Grant, staggered by the majesty of the Brachiosaurus, comments to his colleague Dr Ellie Sattler that this animal couldn’t have lived in a swamp, referring to the re-evaluation of these animals as warm-blooded creatures, closer to the model that the dinosaur-revolution of the ‘60s introduced and opposite to, let's say, modern crocodilians. Although this was partially true, it seems that it is probably incorrect to assume that all dinosaurs were the same when it comes to physiology.
 The sauropod Diplodocus as illustrated by Mary Mason Mitchell for Oliver P. Hay (1910) epitomising the old-fashioned view of dinosaurs.
The sauropod Diplodocus as illustrated by Mary Mason Mitchell for Oliver P. Hay (1910) epitomising the old-fashioned view of dinosaurs.
Our study supports the view that sauropods potentially exhibited a unique physiology, emphasising poikilothermy (higher variation in internal temperature), rather than endothermic homeothermy (maintaining constant body temperature by generating internal heat), and that this was reflected by a distinct palaeobiogeographical signature. A mix of features might have conferred to sauropods a higher adaptation for heat dissipation (compared to modern mammalian analogues), including an avian-like respiratory system, long necks and tails shaping their overall morphology to a higher proportional body surface area, which might have allowed them to avoid overheating in warm environments regardless of their giant sizes. We now know that ornithischians and theropods were characterised (at least in some species) by the presence of insulating integument, which has been suggested as an anatomical correlate of endothermy. No similar integumentary structure has yet been found in sauropods, with evidence of scaly integument with osteoderm-like structures even in fossil embryos, making inferences of feather-like integument in sauropods even less likely. Incubation strategies might also contribute to differences in palaeolatitudinal ranges. Homeothermic theropods probably relied on direct body heat for nest incubation, whereas ornithischians seem to have used microbial decay for incubation. These reproductive strategies might have allowed these groups to be able to nest in polar environments, without having to rely on environmental heat sources. On the other hand, evidence of mound and in-filled hole nesting in sauropods, dependent on solar or geothermic heat sources and more arid climates, might further suggest different thermic preferences even for reproductive strategies.
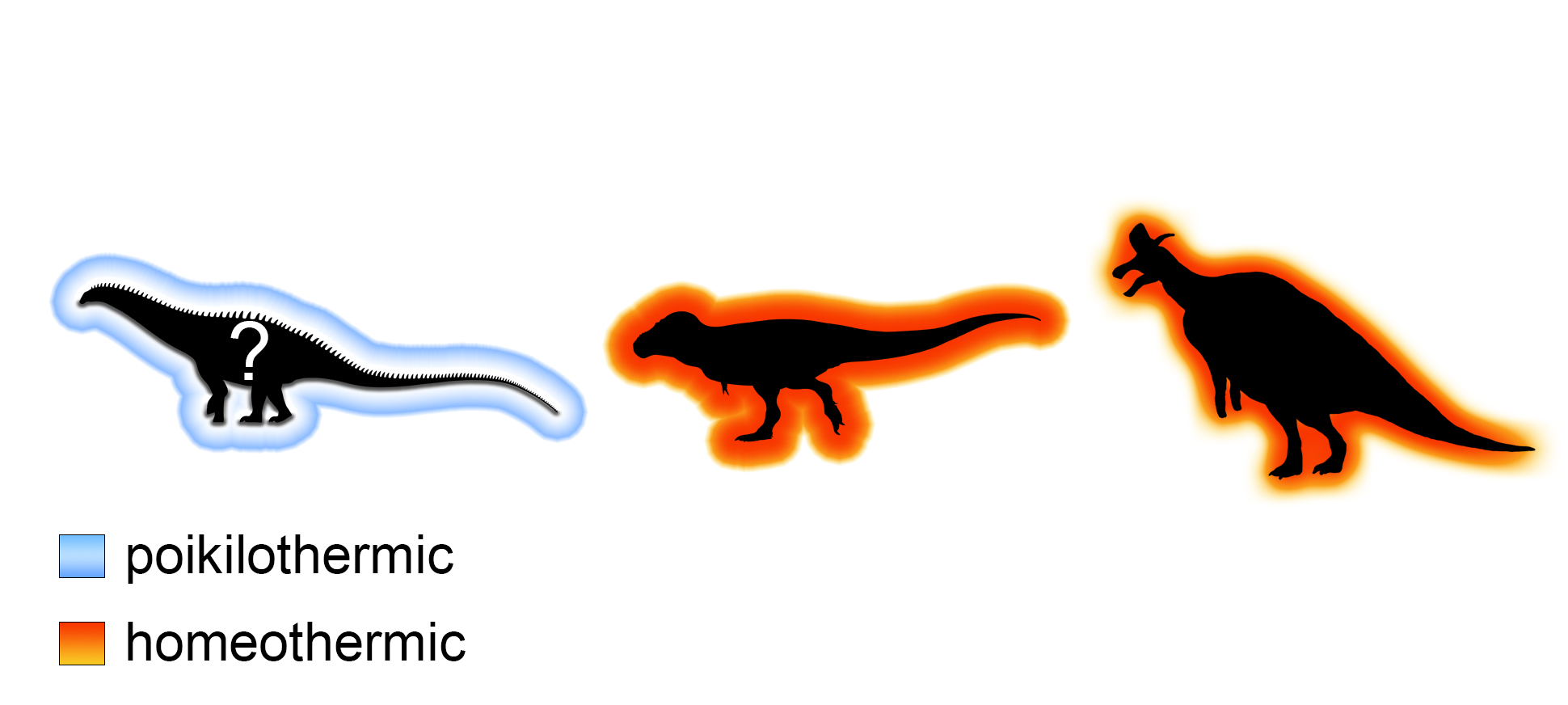
The findings of our study may indicate multifaceted thermal sensitivities in non-avian dinosaurs, which may suggest different thermophysiological strategies, with sauropods emphasizing poikilothermy rather than the endothermic homeothermy likely exhibited by theropods and maybe ornithischians (silhouettes from Phylopic.org).
This unusual dinosaurian biology not only excluded sauropods from cool climatic zones, particularly during colder time intervals, but also regulated the availability of high-latitude dispersal corridors for these animals, including a route from South America to Australasia via Antarctica during the warmest interval of the Cretaceous. This is a unique example of correlating different lines of evidence from the biology and ecology of these extinct animals to broad scale biogeographic patterns. The methodology presented in our study offers a new predictive tool to further constrain biogeographic hypotheses, casting new light on long-standing debates on dinosaur biogeography, palaeobiology, and evolution.

Brontosaurus (likely the most famous sauropod dinosaur) by artist Emiliano Troco, acting as an ecosystem engineer in a warm and vegetated landscape not dissimilar to modern-day savannah type biomes. The animal is basking in a semi-arid open landscape where temperature conditions might have favoured their ecology: as hypothesised in our paper, sauropods might have been an exception in Dinosauria for being solely capable of inhabiting warmer, lower latitude environments, where they might have been more efficient, maybe hinting to a more heterotherm-inclined thermophysiology than other dinosaur groups, i.e. theropods and ornithischians.
I hope you enjoyed the reading, and if you want to know more, the study is published in Current Biology: https://doi.org/10.1016/j.cub.2021.11.061
I am a vertebrate palaeontologist interested in macroecology, mass extinctions and dinosaur palaeobiology. To investigate these topics, I use fossils together with phylogenetic, biogeographic, statistical, and Earth System Modelling tools.


{kind=link}
{kind=link}
{kind=link}