Synergistic antimicrobial specialized metabolites optimized through evolution
Published in Microbiology
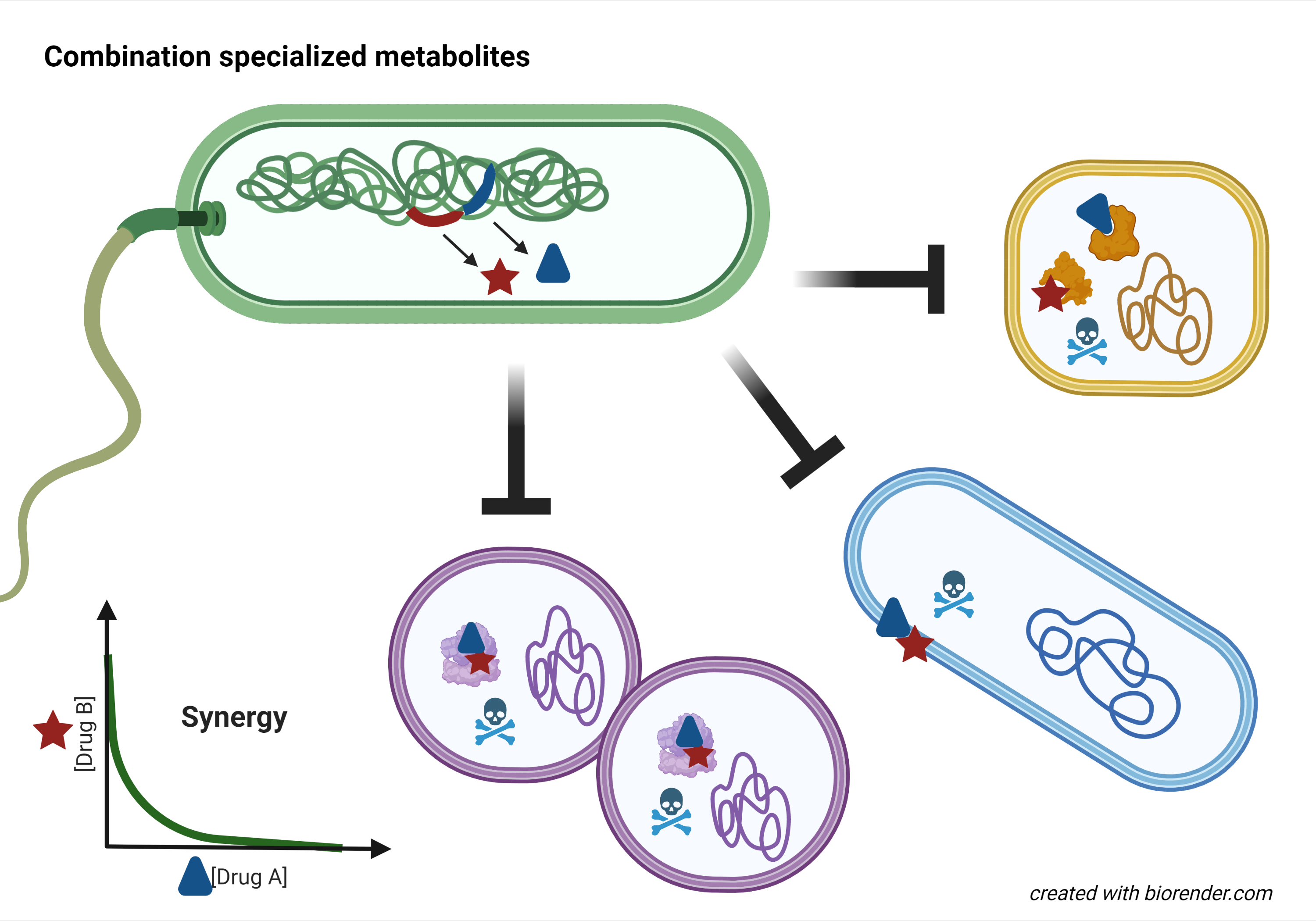
A human gut microbiome Bacillus isolate produces two RiPP peptides that work together to puncture and kill Clostridium difficile1. An amoeba-associated Pseudomonas produces a lipopeptide that enhances the activity of the co-produced tRNA synthetase inhibitor mupirocin, resulting in synergistic inhibition of methicillin-resistant Staphylococcus aureus2. A terrestrial cyanobacterium from a limestone cave uses a branched synthetic pathway to produce two related macrocycles, which together have synergistic antifungal activity3. These are just some of the examples of combination specialized metabolites that captured my imagination as I spent my Covid-19 lockdown digging through the literature. As a pharmacologist and microbiologist, I find these naturally occurring antimicrobial synergistic combinations intriguing. How common are they? How do they work? And can they help us counter the crisis of antimicrobial resistance?
.jpeg)
As I assembled the known cases of these co-produced antimicrobial combinations, I noticed the diversity of the producing bacteria (five phyla represented for sixteen combinations), and the wide variety of ecological niches they had been isolated from. Moreover the antimicrobial metabolites represented a broad range of biosynthetic and chemical classes. Strikingly however, the organizational logic of biosynthetic gene clusters and the regulatory strategy that enabled co-production of these pairs fell into three categories, suggesting either ancient origin or convergent evolution. Altogether I concluded that producing combination metabolites is widespread amongst bacteria.
This makes sense as we continue to learn about the network biology of organisms. So many biological systems integrate multiple signals, have pathways with complex interfaces, and have multiple outputs. For production, genome sequencing and a long history of empirical investigation has taught us that bacteria commonly carry the biosynthetic capacity for dozens of different specialized metabolites. For function, combination antimicrobials mean mechanisms can involve targeting multiple nodes in a pathway for increased efficacy. Bringing these two together, it seems that evolution has selected for the coordinated production and action of multiple specialized metabolites, for increased benefit over a single molecule4.
Despite the consensus that bacterial strains can make many specialized metabolites, and the early precedent of the synergistic streptogramin antimicrobials being discovered in the 1950s5, I believe combination metabolites have been overlooked in research. I identified reports of sixteen distinctive synergistic combinations, whereas there are hundreds of individual metabolites reported with antimicrobial activity. Why is this? Part of the answer may be that typical specialized metabolite identification methods either attempt to identify every metabolite in a bacterial extract or follow bioactivity for single molecule purification. Identifying every metabolite in an extract makes testing the activity of each possible partner combination factorially arduous. When following bioactivity, the less active metabolite of a combination would be lost during purification procedures. From personal experience and communication, it is not uncommon in antimicrobial discovery labs to observe a frustrating loss of bioactivity between crude extract and isolated molecule – perhaps due to loss of a combination partner!
This leads me to the hypothesis that there are many more combination metabolite pairs to be identified. Encouragingly, in recent years genomic data has led to the identification of at least three new synergistic combinations arising from adjacent “superclusters” of biosynthetic genes6, and the identification of new two-component RiPPs7. I also see opportunities to use metabolomics and transcriptomics to identify candidate combination metabolite pairs that are consistently co-produced and co-regulated. An open question is whether combination metabolites are so common as to be produced by almost all bacteria that make specialized metabolites, or whether they will remain a rare and precious jewel in natural product discovery but one which we can develop more effective mining methods to unearth.
The biological role and origins of these combination specialized metabolites is of course a fascinating research direction. Bacterial specialized metabolism has been shaped and selected for over evolutionary time scales to arrive at effective products. The combination examples I review have elegant antimicrobial mechanisms that avoid or neutralize microbial defense systems. In my opinion bacterial combination metabolites are particularly interesting as therapeutic leads for our current crisis of antimicrobial resistance.
Combination therapeutics is now a well-established approach for tackling resistance. Firstly, if the compounds have individual activity and different targets, then two mutations or resistance mechanisms are required which decreases the likelihood of resistance emergence. This is indeed the case for some of the known natural metabolite combinations. The synergistic streptogramins have individual ribosomal inhibition and antibacterial activity, and therefore resistance to the combination is rare both experimentally and clinically8. Secondly, combinations can be used to specifically counter a resistance mechanism. The natural partner metabolite clavulanic acid was the first clinically successful beta-lactamase inhibitor, and has inspired four decades of subsequent development9. Thirdly, potentiator partners that counter innate resistance mechanisms such as the defensive envelopes of pathogens are being actively pursued for clinical development10. One category of the natural synergistic metabolite pairs has this very mechanism. As we uncover more bacterial specialized metabolite combinations, we may discover even more mechanisms that help us tackle resistance.
The complexities and possibilities of combination specialized metabolites fascinate and inspire me. My hope is that the review I’ve written, on synergistic antimicrobial combinations and the mechanisms that drive their production and activity, will support and inspire research and innovation in this most promising area.
Read the review here https://www.nature.com/articles/s41564-021-00952-6
1 Rea, M. C. et al. Thuricin CD, a posttranslationally modified bacteriocin with a narrow spectrum of activity against Clostridium difficile. Proc Natl Acad Sci U S A 107, 9352-9357, doi:10.1073/pnas.0913554107 (2010).
2 Arp, J. et al. Synergistic activity of cosecreted natural products from amoebae-associated bacteria. Proc Natl Acad Sci U S A 115, 3758-3763, doi:10.1073/pnas.1721790115 (2018).
3 Heinilä, L. M. P. et al. Shared PKS module in biosynthesis of synergistic laxaphycins. Front Microbiol 11, 578878, doi:10.3389/fmicb.2020.578878 (2020).
4 Challis, G. L. & Hopwood, D. A. Synergy and contingency as driving forces for the evolution of multiple secondary metabolite production by Streptomyces species. Proc Natl Acad Sci U S A 100 Suppl 2, 14555-14561, doi:10.1073/pnas.1934677100 (2003).
5 Charney, J., Fisher, W. P., Curran, C., Machlowitz, R. A. & Tytell, A. A. Streptogramin, a new antibiotic. Antibiot Chemother (Northfield) 3, 1283-1286 (1953).
6 Alanjary, M. & Medema, M. H. Mining bacterial genomes to reveal secret synergy. J Biol Chem 293, 19996-19997, doi:10.1074/jbc.H118.006669 (2018).
7 Walker, M. C. et al. Precursor peptide-targeted mining of more than one hundred thousand genomes expands the lanthipeptide natural product family. BMC Genomics 21, 387, doi:10.1186/s12864-020-06785-7 (2020).
8 Reissier, S. & Cattoir, V. Streptogramins for the treatment of infections caused by Gram-positive pathogens. Expert Rev Anti Infect Ther, 1-13, doi:10.1080/14787210.2021.1834851 (2020).
9 Reading, C. & Cole, M. Clavulanic acid: a beta-lactamase-inhiting beta-lactam from Streptomyces clavuligerus. Antimicrob Agents Chemother 11, 852-857, doi:10.1128/aac.11.5.852 (1977).
10 Tyers, M. & Wright, G. D. Drug combinations: a strategy to extend the life of antibiotics in the 21st century. Nat Rev Microbiol 17, 141-155, doi:10.1038/s41579-018-0141-x (2019).
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in