The Tethering Complex Revisited: Discovery of New Complexes Regulating Vesicle Fusion in Endocytic Pathways
Published in Cell & Molecular Biology
The tethering complexes HOPS and CORVET are multi-protein complexes that regulate the fusion of vesicles such as endosomes, autophagosomes, and lysosomes in endocytosis and autophagy pathways. The importance of these complexes in intracellular vesicular trafficking has been elucidated by many research groups. In the present study, we have shown that there are hybrid types of complexes formed by different combinations of subunits which constitute HOPS and CORVET, and that this complex functions in a certain type of endocytosis.
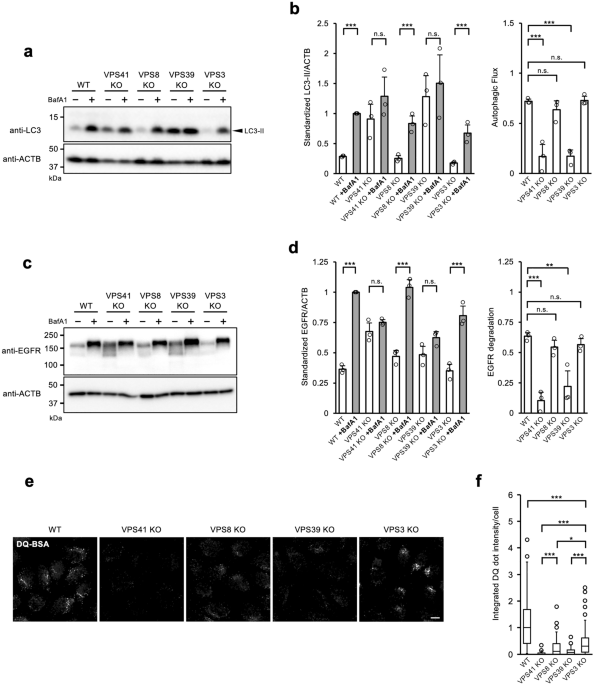
Our study was initiated by the identification of several mucopolysaccharidosis-like patients. Mucopolysaccharidosis is an inherited metabolic disorder caused by a deficiency of lysosomal enzymes such as α-L- iduronidase or iduronate-2-sulfatase. However, all relevant enzyme activities in these patients were normal. Whole exome analysis revealed that these patients had a common mutation (p.R498W) in the VPS33A gene, one of the subunits of the tethering complexes. As it is a mutation involved in HOPS and CORVET, we suspected that it affects autophagy and/or endocytosis. We decided to generate a comprehensive series of knock-out cells for each subunit gene of HOPS and CORVET in HeLa cells and investigated their effects on autophagy and endocytosis in detail.
When assessing the effects of these gene deletions on macropinocytosis, we noticed that VPS8 KO and VPS3 KO cells behaved differently. Since both VPS8 and VPS3 are subunits constituting CORVET, comparable results should be observed if they reflect the consequences of CORVET loss. Thus, we hypothesized that a previously unknown complex may exist. We labeled each subunit with a different tag and investigated what kind of complexes were formed by Tag-immunoprecipitation, and found that there were hybrid complexes in which the combinations between VPS41/VPS8 and VPS39/VPS3 were swapped (Fig. 1). We, therefore tentatively refer to these new complexes as Hybrid-A and Hybrid-B, nevertheless we are thinking to give them more suitable names in near future. The existence of the hybrid-type complexes was further thoroughly verified by single-subunit pull-down, endogenous complex IP, and mass spectrometry.
, and the newly-identified complexes (right half).")
Many may wonder why the Hybrid type of complexes have not been found until now, albeit so many researchers have been studying the tethering complexes. In fact, the existence of this complex was previously suggested in yeast. Peplowska et al. found complexes similar to ours and reported them as iCORVET and iHOPS (Peplowska et al., Dev Cell., 12, 2007). We were also inspired by this study. By overexpressing each subunit of the tethering complex and immunoprecipitating it, we got a glimpse of the existence of a hybrid-type complex in the very early study, but the results were not easily reproduced. Far from it, even known complexes, HOPS and CORVET were just barely able to be detected in immunoprecipitation. After repeated investigations, we found that even very low concentrations of detergent destabilized these complexes. Therefore, we changed to using a syringe disruption method for the preparation of cell lysates without detergent, and then all complexes became to be detected stably. We assume that many people use detergents in immunoprecipitation, and this may be one of the reasons why hybrid-type complexes have not been noticed so far. Even when detected under optimized conditions, the amount of Hybrid-B is very low compared to the other complexes. As far as examined with the fixed viewpoints of only HOPS and CORVET, no one would have noticed the presence of the Hybrid-type complexes.
In the report by Peplowska et al., the hybrid-type complexes were regarded as intermediates in the formation process of HOPS or CORVET. Since the four complexes share several subunits, it is not easy to analyze the respective functions of the complexes. In this study, by comparing and analyzing single KO cells and double KO cells of the subunits other than the class C core moiety, we succeeded in shedding light on the function of the Hybrid-type complex in endocytosis. While receptor-mediated endocytosis (RME) can be driven by HOPS alone, we showed that Hybrid-B contributes significantly to at least liquid-phase endocytosis such as macropinocytosis (Fig. 2). This means that it cannot rule out the possibility that endocytosis, which has been recognized to be driven by HOPS and CORVET, may include the Hybrid-B function. On the other hand, Hybrid-A is more abundant than Hybrid-B or HOPS, but we could not figure out a specific function of this complex with statistically significant data. We are looking forward to future studies on the Hybrid-A function.
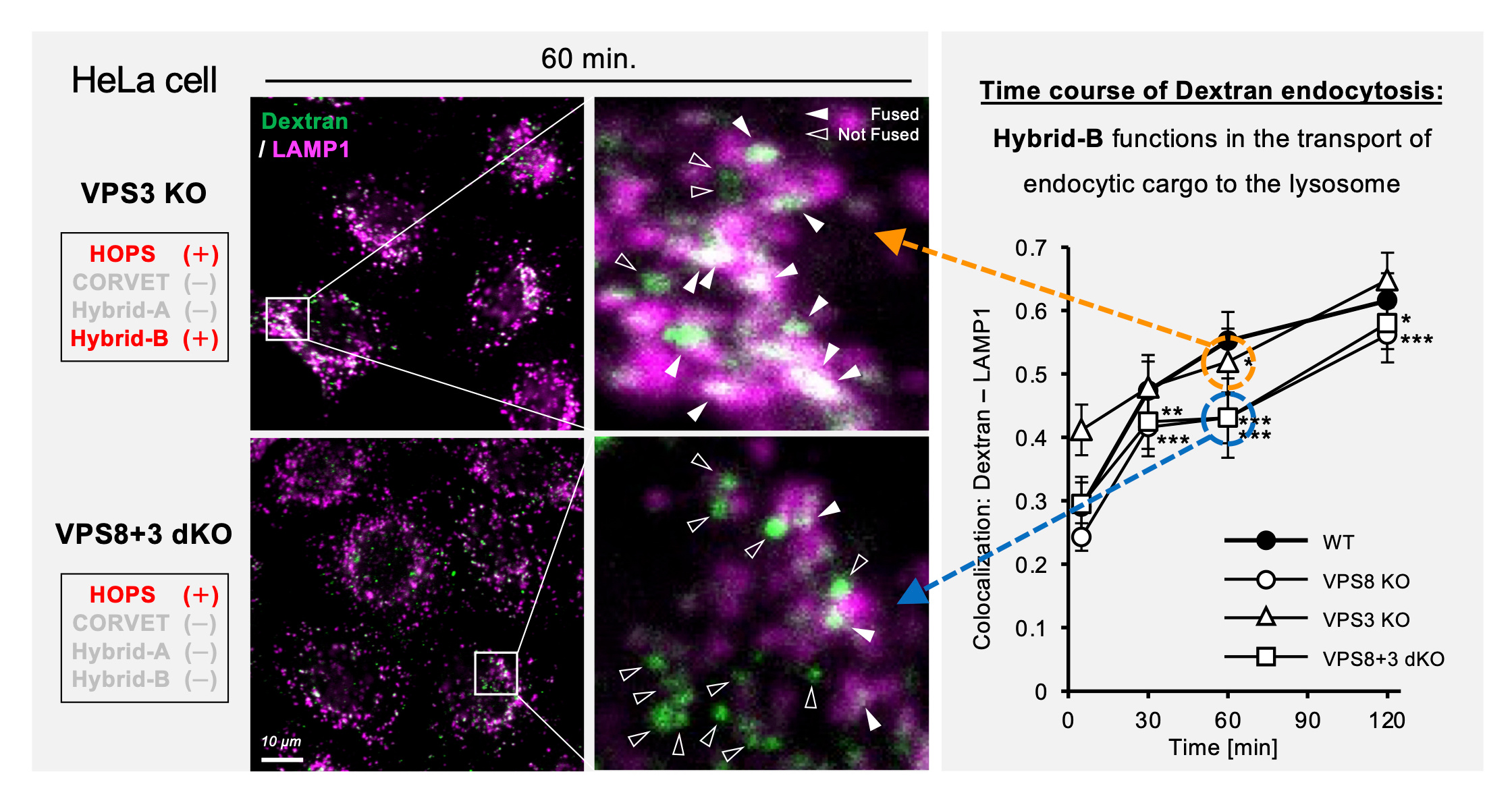
Figure 2. Hybrid-B functions in macropinocytosis. The trafficking of fluorescence-labeled dextran to lysosome was compared between VPS3 KO and VPS8+3 double KO (dKO) cells at the time point of 60 minutes after pulse (microscopic images). The right panels are enlarged pictures of the inset in the left panels. The time course of dextran transport to lysosome was evaluated by Pearson’s correlation coefficient between dextran and LAMP1 signals (graph). The corresponding data to VPS3 KO (orange) and VPS8+3 dKO (Blue) were marked and connected with circles and arrows to the microscopic images.
HOPS is believed to be involved in the fusion between endosomes and lysosomes at the end stage of endocytosis. In fact, our studies have confirmed that endocytosed substrates cannot reach lysosomes in the KO cells lacking HOPS. Interestingly, in the HOPS-deficient cells, both EGF; a substrate for RME, and dextran; a substrate for macropinocytosis, were transported to the vicinity of lysosomes after uptake, however, the majority of these substrates appeared to separately reside in different vesicles (Title background). The general endocytosis model that has been considered so far seems to suggest that substrates are first collected in early endosomes and are transported to lysosomes via a common route from there. Still, our study raises the possibility that they are transported through a different route until just before fusion with lysosomes. As mentioned above, the fact that Hybrid-B does not impact RME but does function in liquid-phase endocytosis may also support the possibility that each route is differentially regulated. The endocytosis pathway may be a more sophisticated logistic system under elaborate control than we have imagined. Our findings suggest that there is still much to be elucidated in endocytosis. We will continue to elucidate the function of hybrid-type complexes in intracellular vesicular trafficking.
Follow the Topic
-
Scientific Reports
An open access journal publishing original research from across all areas of the natural sciences, psychology, medicine and engineering.

Related Collections
With Collections, you can get published faster and increase your visibility.
Infectious disease diagnostics
Publishing Model: Open Access
Deadline: Sep 23, 2026
AI in Education
Publishing Model: Open Access
Deadline: Oct 09, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in