The wild side of Arabidopsis thaliana and its associated root microbiota
Published in Ecology & Evolution


Plants growing in their natural habitats are constantly facing environmental stresses and responding to a myriad of cues. Furthermore, these plants are completely surrounded and colonized by a huge variety of microbes, which include bacteria and filamentous eukaryotes such as fungi and oomycetes (Turner et al. 2013, Muller et al. 2016). In the same way that their host plants are affected by their surroundings, the plant-associated microbiota is also influenced by environmental conditions. However, it remains unclear to which extent the local environment affects the composition of the plant root microbiota across large spatial scales.
In 2013, my PhD mentor Stéphane Hacquard (Max Planck Institute for Plant Breeding Research – MPIPZ-, Cologne, Germany) decided to get out of the lab and collect the model plant species Arabidopsis thaliana growing in natural conditions. The project started with few natural plant populations near the MPIPZ, which were initially used to establish comprehensive microbial culture collections from plant roots, with which we then tested the relevance of microbe-microbe interactions for plant survival under laboratory conditions (Durán et al. 2018). When I joined the lab in 2014 to pursue my PhD, we decided to set-up a more comprehensive survey of the A. thaliana root microbiota, conclusions of which are now published in the current issue of Nature Ecology and Evolution (Thiergart et al. 2019).
First of all, we had a few basic questions: 1) What is the Arabidopsis thaliana root associated microbiota composed of? 2) Is it conserved across different sites? 3) Is it similar to the microbiota of other co-occurring plants? 4) What environmental factors affect this microbiota? By profiling root-associated microbial communities associated with our favorite model plant across many different soil conditions and climates, we expected to gain deeper insights into how the local environment affects microbial community composition at the root interface. In order to tackle all these questions, we decided to select several A. thaliana populations naturally occurring across a latitudinal gradient in Europe. For that, we teamed-up with European experts on A. thaliana populations, including Carlos Alonso Blanco (CNB, Madrid, Spain), Fabrice Roux (INRA Toulouse, France), Jon Ågren (University of Uppsala, Sweden) and Eric Kemen (who, at the time, was at the MPIPZ, Germany). Once we had them all on board, we designed the sampling so that we would harvest A. thaliana plants (their expertise being instrumental on plant identification in the wild, more about this later…) at the same phenological stage (flowering plants) and surrounding soil in each site, and then transport it back to the MPIPZ. In this way, we would have four harvesting excursions (one per country) across four months (February to May) over three consecutive years (spring 2015, 2016 and 2017).

Stéphane and I teamed up with Samuel Kroll and Alfredo Mari, two PhD students working in the group of Eric Kemen at the time, who also took advantage of the trips to address their own research questions (Figure 1). After all the preparatory work and gearing up for the unknown, the four of us drove to each of the initially 16 locations and met up with our local collaborators, who would show us where A. thaliana grows and help us to identify them. This last point, although it may seem trivial, turned out to be of extreme importance. One would think that plant scientists who had been working on A. thaliana for a number of years would easily recognize their model plant in the wild. Well, you would be surprised. And we were too: when growing A. thaliana in the greenhouse, we very often get big, happy plants with huge rosettes; however, this is very far away from what these plants look like in nature. In our very first harvesting excursion, we had a very hard time to spot these tiny plants and even sometimes to differentiate them from closely related plant species such as Draba verna (Figure 2). After collecting plants from so many sites over several consecutive years, I now can see immediately the difference, but it was definitively not so clear at the time…

After many kilometers and many hours looking for tiny, often hidden and phenotypically very different plants, we managed to collect 174 individuals. Roots of these healthy plants were fractionated into four microbial niches in a distance gradient from the root interior: Soil, Rhizosphere, Rhizoplane and Root (Figure 3). Then, after many, many, many PCRs and several MiSeq runs, we obtained our first sneak peak of the data (almost a year after the project started!). At this point, Thorsten Thiergart (co-first author of this work) joined the lab and made some crucial advances in data processing to allow simultaneous analysis of bacterial, fungal and oomycetal communities from each of the root fractions.

This first data analysis told us many things, but one of the most striking observations was the robustness of the composition of bacterial communities in root tissues. Despite large geographical distances and different soil conditions and climate between sites, the composition of bacterial communities in the root compartment was more similar across sites than the surrounding soil communities. This similarity in root microbiota composition was not as pronounced for fungal and oomycetal communities, although a host-filtering effect can also be observed for these microbial groups (Figure 4). Based on these first observations, we were excited to pursue the harvesting campaign for two more years. This successive sampling allowed us to assess the robustness of these community patterns over time, and to determine the extent to which year-to-year variation in environmental conditions affect microbial community composition.
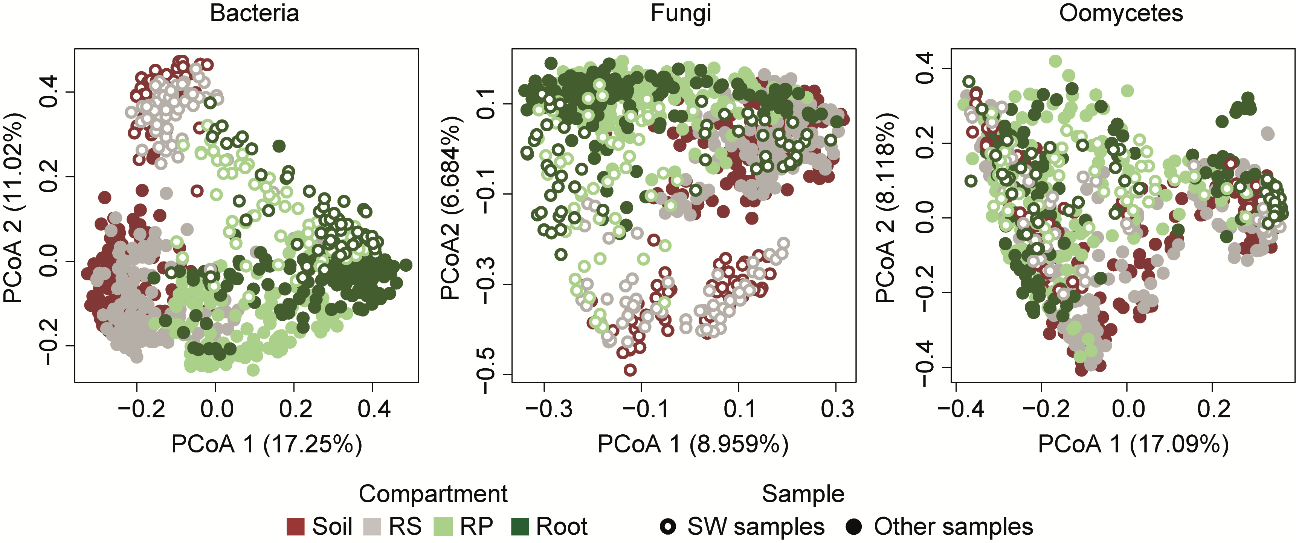
Results from these additional harvesting campaigns not only strengthened our initial observations, but also allowed us to draw one more conclusion: community composition of filamentous eukaryotic communities are highly variable across years, contrasting with the composition of bacterial communities that remains remarkably stable at the root interface (Figure 4). Furthermore, analysis of soil properties and correlation with microbial abundance profiles confirmed that the variables “pH” and “iron” extensively explain microbial community variation in soil. However, these parameters only poorly explain root microbiota differentiation across sites. Most remarkably, we were able to identify root microbiota members that were consistently found in A. thaliana roots across sites and years, which will allow us to rationally design synthetic microbial communities for reductionist approaches in the lab.

Although we identified conserved signatures in root microbiota composition across European sites, we also observed that the local environment (i.e. the factor “site”) shaped the root microbiota. However, the “site” did not tell us which of the many site-specific factors (such as soil conditions, climate or host genotype) was primarily driving the observed variation. During this time, one of our collaborators, Jon Ågren, had a common-garden experiment running in one of our Swedish locations as well as at an Italian location close to Rome. In this experiment, Jon was initially testing the extent to which the two geographically widely separated A. thaliana populations in Sweden and Italy are adapted to their local soil conditions and local climate. In this field transplant, the Swedish and Italian A. thaliana genotypes were swapped between the two locations and were grown in both Swedish and Italian soils at each site (Figure 5). This actually turned out to be exactly what we needed to disentangle the previously overlapping “site” factors in our European transect experiment: host genotype, soil properties and climate. Therefore, we included the Italian site in our microbiota transect survey and also harvested root and soil samples from plants grown at both sites in both soils. Furthermore, our Swedish collaborators scored A. thaliana fitness, another parameter that we were previously not able to obtain.
Analysis of these samples showed that the make-up of bacterial communities in plant roots remained strongly dependent on host compartment and soil conditions, whereas the identity of eukaryotic members was largely specified by changes in climatic conditions. Surprisingly, although significant, host genotype was a relatively minor factor affecting microbial community composition. Furthermore, the two host genotypes were found to produce more fruits when grown in their local soil and climate, providing evidence for strong local adaptation between these two plant populations in Northern and Southern Europe.
In conclusion, we comprehensively described bacterial, fungal and oomycetal communities associating with roots of A. thaliana in wild populations and characterized how environmental cues can affect their distribution across large spatial scales. Secondly, our common-garden experiment allowed us to uncouple the “site” effect into climate, soil conditions and host genotype.
From this experience, I will take with me the adventures involved in travelling to different countries and to different labs in the course of my PhD. I could also see how stimulating it can be to interact with researchers from different fields and with different backgrounds, and also to see how things work in different institutions. Of course, it was also a plus to have some time outside of the lab getting some fresh air, despite the climatic conditions (just picture us digging up plants in freezing-cold temperatures in Sweden, or plant-hunting in a surprisingly rainy Spain). Also, I will cherish all the botanical knowledge I did not expect to gain at the beginning of my PhD as well as all the bioinformatics insights, which I did not anticipate at all. And last but not least, after this experience, we are very happy to now share with the scientific community our fruitful collaborative project in this issue of Nature Ecology and Evolution. We hope you enjoy it!
References
Turner, T. R., James, E. K., & Poole, P. S. (2013). The plant microbiome. Genome biology, 14(6), 209.
Müller, D. B., Vogel, C., Bai, Y., & Vorholt, J. A. (2016). The plant microbiota: systems-level insights and perspectives. Annual review of genetics, 50, 211-234.
Durán, P. et al. Microbial Interkingdom Interactions in Roots Promote Arabidopsis Survival. Cell 175, 973–983.e14 (2018).
Thiergart, T., Durán, P., Ellis, T. et al. Root microbiota assembly and adaptive differentiation among European Arabidopsis populations. Nat Ecol Evol 4, 122–131 (2019)
Follow the Topic
-
Nature Ecology & Evolution
This journal is interested in the full spectrum of ecological and evolutionary biology, encompassing approaches at the molecular, organismal, population, community and ecosystem levels, as well as relevant parts of the social sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Understanding species redistributions under global climate change
Publishing Model: Hybrid
Deadline: Dec 31, 2026


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in