Alternative history of life on Earth: complex life evolving from giant sulfur bacteria
Published in Ecology & Evolution and Microbiology

What if Asgard archaea never emerge, and an archaeon never met a bacterial endosymbiont? I have bad news for you in this case — we would never exist, as well as other eukaryotes. The Great Oxidation Event would still occur due to cyanobacteria, and the atmosphere would still be oxidizing and full of oxygen. But mitochondria would never arise and never kick-start the process of eukaryogenesis.
Does this mean that the world would be totally devoid of complex life, such as compartmentalized cells and multicellular life? It could seem so because prokaryotes generally have low potential for intracellular complexity (the only bacteria with the separate “nuclei” are planctomycetes) and multicellularity (when DNA is not separated from the cytoplasm, gene regulatory networks are generally less elaborate). And still there is a group of bacteria that would have potential to save the world in these circumstances. They are called giant sulfur bacteria.
These bacteria have a peculiar biochemical specialization: they thrive in the water bodies where plenty of hydrogen sulfide is available. They absorb it and oxidize sulfide to sulfate, thus obtaining energy from the oxidation of inorganic compounds (this way of living is called chemolithotrophy, in literal translation — “using the chemicals from stones as food”). To perform this process, they need an oxidizer – and they normally use oxygen like we do for our organic food (we are organoheterotrophs, but we use the same oxidizer).
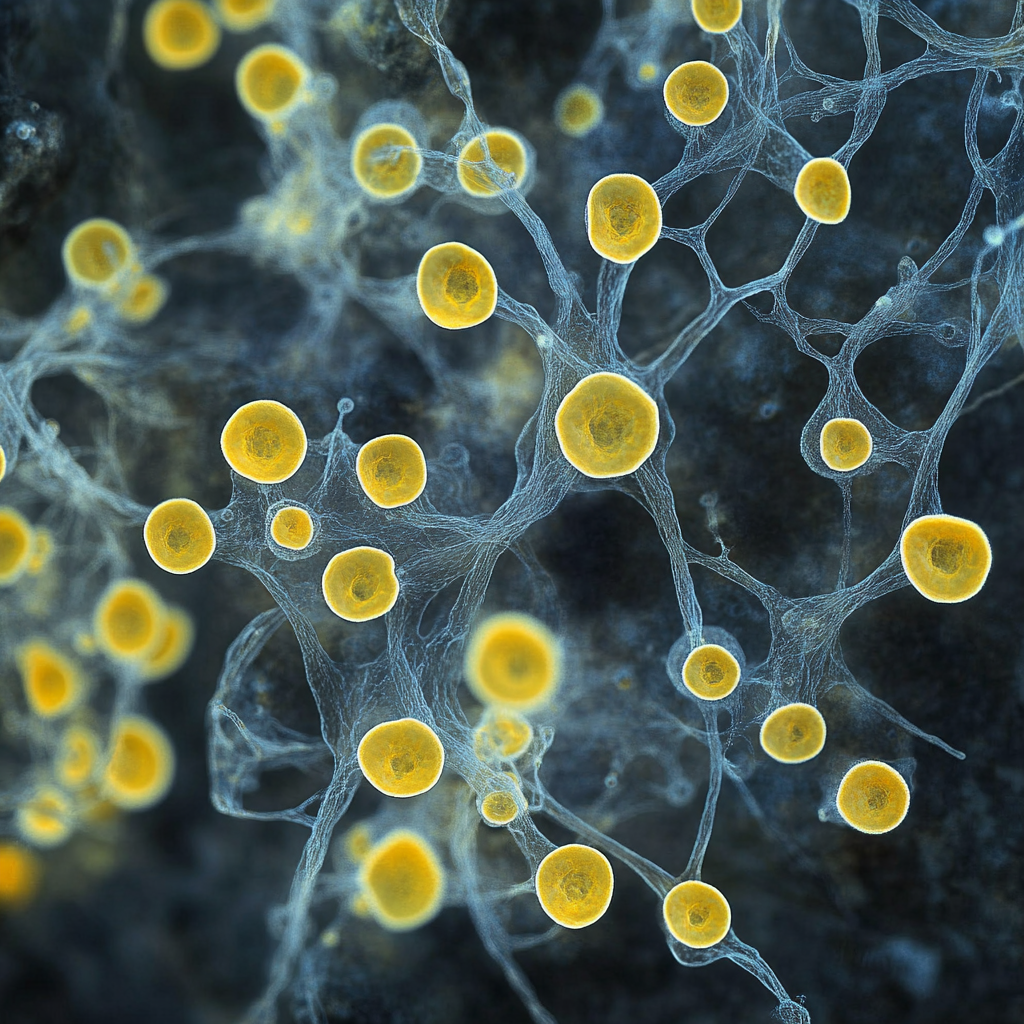
But then, strange things begin. Hydrogen sulfide normally is found only in deep layers of some water bodies, being supplied there by geological sources (oil gas, natural gas, volcanic gases) or by organic matter decomposition. In contrast, oxygen is located in the upper layers, being supplied from the atmosphere by diffusion. This creates a top-down chemical gradient of oxygen and sulfide — while the bacteria need both compounds.
This creates a pressure selection towards being long enough to span the pools of oxygen and sulfide and use them both (Fig. 1). This raises quite different strategies among giant sulfur bacteria to achieve extremely large size on a prokaryotic scale.

Masters of complexity
Let’s imagine a typical prokaryotic cell. It has almost nothing more than a membrane and is a simple sphere in the terms of topology. It has a fundamental restriction on their size: its energy supply depends on its membrane area. Almost all living cells generate the major energy molecule — ATP — through membrane-bound ATP synthases using transmembrane proton gradients. This design is a possible relic from earlier life, the proto-cell era, when the mineral proto-cells had an abiogenic proton gradient on their membranes (as Nick Lane explains in his lecture for the Royal Society of Biology). In contrast, energy consumption depends on the cell volume, because energy-consuming enzymes are scattered across the entire cell volume.
This gives birth to energy supply-consumption discrepancy: when the length of a bacterium increases, its energy supply grows proportionally to the square of its linear dimensions, and its energy consumption grows proportionally to the cube of these dimensions. This restricts the size of a typical bacterium, and this is the cause why bacteria are too small to be seen by a naked eye. But giant sulfur bacteria really need to be giant and extremely long — this is the matter of their survival. But how to overcome this mathematical restriction? There is a solution: to increase complexity.
For a small sphere-formed cell, there are two main strategies to grow bigger and longer (Fig. 2). First one can be called "multicellularization": joining many simple sphere-like cells together, until their total length meets the selection requirement. This implies increasing the number of the cells — the simplest metric of complexity. This consideration is confirmed by several evolutionary experiments: unicellular organisms evolved multicellular structures under the positive selection for larger size [1, 2].
The second strategy can be called "eukaryotization": putting a lot of simple sphere-like cells inside one big sphere-like cell. This also implies growing the number of these "mini-cells". This was a real innovation of eukaryotes — not the nucleus itself. And this is the reason why planctomycetes never rival eukaryotes by complexity. In the both cases, positive selection towards size in prokaryotes means the positive selection towards complexity.

What is unique about giant sulfur bacteria? The fact that — being the members of only one order, Thiotrichales, — they are extremely diverse in the terms of ways to complexity. Some bacteria, like Beggiatoa, keep track of multicellularization, but others, like recently discovered Thiomargarita magnifica, explores the way of eukaryotization. This unprecedented diversity has confused me in the course of my own research — and the puzzle went together only with the discovery of Thiomargarita magnifica. (I told this story in one of my previous posts and in an arfticle in Biochemistry (Moscow) [3]).
Thiomargarita magnifica is not a true eukaryote, but has a lot of prerequisites to the eukaryotic level of complexity. Coexistence of multicellular and eukaryote-like forms within one order shows that the necessity to bridge together the substrate pools produces really enormous selection pressure towards size and complexity. It also suggests that giant sulfur bacteria could develop both cells with eukaryotic level of complexity and a plenty of multicellular forms — and create living landscape of our planet if true "chimeric" eukaryotes never arise.
Look at this giant sulfur world
Ok, let's imagine our apocalyptic world. About 2 million years ago, a time traveler tried to cure his wound with a sulfanilamide powder and accidentally spilled the antimicrobial powder onto the last common ancestor of all Asgard archaea. Archaea are susceptible to sulfanilamide like bacteria (it makes them devoid of folic acid the same way), so the archaeon dies, and Asgards never emerge. This is a time-travel paradox worse than a killed grandfather. But giant sulfur bacteria could still give our planet a chance be something more than just a boring barren landscape.
How would this world look like? I tried to imagine using AI services as assistants. I prompted two AIs: a well-known Midjourney and a built-in generative AI of MindTheGraph service.
What to start with? I assumed that long bodies of giant sulfur bacteria could give rise to a wide variety of long branched non-motile multicellular creatures. But this is also one of the most frequently emerging multicellularity types in our normal world [4] and found, for example, in plants and fungi. But some giant sulfur bacteria, like Beggiatoa, accumulate elemental sulfur inside their cells as an intermediate product — so I prompted generative AIs to draw fungi-like or plant-like structures accumulating sulfur inside their cells.

In the "giant sulfur world", we would see mycelial organisms highly similar to modern fungi, but with an extra peculiarity as clumps of sulfur (Fig. 4,5). Their long mycelium would bridge sulfide-rich abysses and oxygen-rich air (cyanobacteria are still alive in this universe) without any problems. Maybe, they even could form fruiting bodies — not on lush, green and wet forest floor, but on barren ground with the only nutrient — hydrogen sulfide — deeply below (Fig. 6).



Even giant plant-like organisms could arise (Fig. 7). Yes, they would not photosynthesize (poor cyanobacteria, you would need to be a devil for work!), but their tree-like appearance and branched roots should enable them to have access to both aerial oxygen and underground hydrogen sulfide. We should see the forests of yellow trees with strange structures instead of leaves... and the planet would live in eternal meadow autumn.

What about animal-like forms? On one hand, existing at the sulfide-oxygen boundary puts some restrictions on the life forms — and multicellular organisms with motile cells seem unexpected. On the other hand, animal multicellularity is an unusual type even in our world, and its emergence has been a kind of a "black swan event" [4, 5]. Thus, since it emerged here, it had a chance to emerge there. After all, some giant sulfur bacteria could use the both their substrates due to their motility, literally holding their breath to accumulate one substrate, then move and use another one [6]. And in our hypothetical world, this ability might be developed further. We could hope, at least, to see aquatic coral-like and meduza-like animals: inevitably with yellow parts because of sulfur, but so familiar to us (Fig. 8, Fig. 9).


Thus, all forms of clonal multicellularity would have the chance to emerge in this world. I just cannot find any plausible scenario to build aggregative forms (such as dictyostelid slime moulds) in these conditions. But then, if they specialized on oxidizing excessive elemental sulfur on the ground... we could see something like Fig. 10.

Take-home messages
This visual imaginary experiment (with the aid of artificial intelligence) is more than a science entertainment. First of all, it emphasizes and demonstrates the potential of giant sulfur bacteria to develop complex structures and the dependence of complexity on biochemical conditions.
But it also could help to answer an everlasting question of modern evolutionary biology. Were the emergence of eukaryotes a determined, or rather an accidental event?
Mounting evidence suggests that eukaryogenesis could be kick-started by a syntrophic association between an Asgard archaeon and a sulfate-reducing bacterium [7, 8, 9]. Such syntrophic associations between archaea and sulfate-reducing bacteria are relatively common, but a recent discovery shows that they could also tend to form complex structures and interactions. Direct conductometry confirmed that anaerobic methanotrophic archaea form a living electrical network together with sulfate-reducing bacteria (Fig. 11) for interspecies electron transfer and working on the same reaction together [10].

This is a clear sign that archea-bacterial symbioses — especially of this type — have powerful biochemical preconditions for complexity, like giant sulfur bacteria do. But it is a game of chance which form would this complexity take.
For us, eukaryotes are only a specific type of complexity, because we have not seen any other reference for "eukaryote". By this thought experiment, I tried to look on the problem through a broader lens and show, that sometimes all we need to get insights on a scientific problem is to engage our imagination.
And please, treat the environment sustainably, including microbes. Let's save current multicellular life, to have no concerns about saving last common ancestors for the next one.
Update from May 19, 2026
This article (its Russian translated version published by Biomolekula magazine) took second place in BioMolText-2025/2026 contest.
The inspiring feedback from colleagues also encouraged me to write a preprint [11] based on this article. It includes less AI content, but summarizes key scientific points and provides important addition — including considerations on cable bacteria.
References
- W.C. Ratcliff, R.F. Denison, M. Borrello, & M. Travisano (2012) Experimental evolution of multicellularity. Proc. Natl. Acad. Sci. U.S.A., 109 (5), 1595-1600, https://doi.org/10.1073/pnas.1115323109
- Herron, M.D., Borin, J.M., Boswell, J.C. et al. (2019) De novo origins of multicellularity in response to predation. Sci. Rep., 9, 2328. https://doi.org/10.1038/s41598-019-39558-8
- Kurakin, G. (2023) Lipoxygenase in a Giant Sulfur Bacterium: An Evolutionary Solution for Size and Complexity? Biochemistry (Moscow), 88, 842–845. https://doi.org/10.1134/S0006297923060111
- Brunet T, King N. (2017) The Origin of Animal Multicellularity and Cell Differentiation. Dev. Cell, 43 (2):124-140. https://doi.org/10.1016/j.devcel.2017.09.016
- Ruiz-Trillo, I., Kin, K., & Casacuberta, E. (2023)The origin of metazoan multicellularity: a potential microbial black swan event. Annual Review of Microbiology, 77 (1), 499-516. https://doi.org/10.1146/annurev-micro-032421-120023
- Schulz, H. N. (2002) Thiomargarita namibiensis: Giant microbe holding its breath, ASM News, 68, 122-127, https://doi.org/10.1128/AEM.68.11.5746-5749.2002
- Purificación López-García, David Moreira. (2020) The Syntrophy hypothesis for the origin of eukaryotes revisited. Nature Microbiology, 5 (5) 655–667. https://doi.org/10.1038/s41564-020-0710-4
- Purificación López-García, David Moreira. (2020) Cultured Asgard archaea shed light on eukaryogenesis. Cell, 181 (2), 232–235. https://doi.org/10.1016/j.cell.2020.03.058
- Mart Krupovic, Valerian V. Dolja & Eugene V. Koonin. (2023) The virome of the last eukaryotic common ancestor and eukaryogenesis. Nature Microbiology, https://doi.org/10.1038/s41564-023-01378-y
- Hang Yu, Shuai Xu, Yamini Jangir, Gunter Wegener, Victoria J. Orphan, Mohamed Y. El-Naggar. Redox conduction facilitates direct interspecies electron transport in anaerobic methanotrophic consortia. Science Advances, https://doi.org/10.1126/sciadv.adw4289
- Kurakin, G. (2026). World Without Eukaryotes: Alternative Evolution of Complex Life from Giant Sulfur Bacteria. Preprints, 202605.1092. https://doi.org/10.20944/preprints202605.1092.v1
Biologist focused on bioinformatics, multicellularity evolution, and cross-kingdom pathogenicity.
Laboratory medicine doctor by training.
Member of the Royal Society of Biology (RSB) and International Society of Computational Biology (ISCB).
Science communicator, regular contributor of The Skeptic magazine (skeptic.org.uk).
Digital artist by hobby (see my website).
Living in Tver', Russia.





Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in