Asymmetric chromatin capture by the MTOC and nuclear envelopes separate chromosomes in fused cells in vivo
Published in Healthcare & Nursing

Cell-cell fusion
Cell-cell fusion is a highly regulated process that occurs in normal physiological conditions. Examples include the fusion of the sperm with the egg during fertilization; the fusion of trophoblasts during placenta formation; and the fusion of myoblasts during muscle formation1. Cell fusion has also been proposed to be involved in cancer initiation and progression. It has been reported that the hybrids produced by fusion in vitro or in vivo display aneuploidy and show higher metastatic potential2,11. However, the consequences of cell fusion on cell cycle progression, spindle assembly, chromosome segregation, cell fate, and other basic cell biological processes are incompletely understood. We were particularly interested in understanding the formation of dual spindles and the chromosome separation that has been reported in fused cells, such as during the first division after fertilization in certain insect species, arthropods, and mouse zygotes; and in hybrid cells derived from somatic cell-cell fusion3–6.
Method
We developed a cell-cell fusion assay, where a 532nm pulsed laser was used to induce a small lesion at the interface between Drosophila neural stem cells (neuroblasts (NBs)) with differentiating ganglion mother cells (GMCs) in the intact fly larval brain. This way we were able to generate hybrid cells in vivo between molecularly distinct cell types and ask how these epigenetically distinct chromosomes will be recognized, separated, and segregated in hybrid cells.
Mitosis of the NB-GMC hybrid
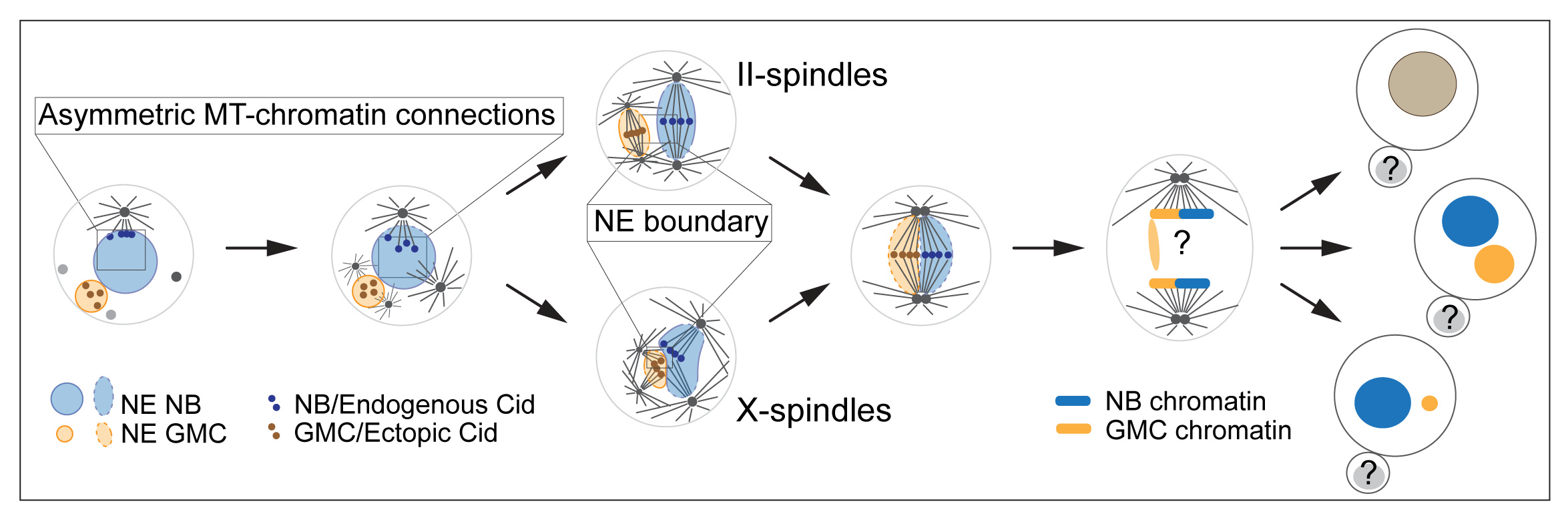
We found that NB-GMC hybrid cells keep the neuroblast chromatin separate from the GMC chromatin and align them independently at the metaphase plate. These hybrid cells usually contain two neuroblast- and two GMC-derived centrosomes, which form spindles that are positioned next to each other during metaphase, thereby congressing the neuroblast- and GMC-derived chromatin at the metaphase plate. The NB and GMC chromatin can initiate segregation autonomously suggesting that the dual spindles in the hybrid cell can operate independently. As stated above, we observed that the NB and GMC chromatin remain separated at the metaphase plate and maybe through anaphase.
How is the separation of the NB and GMC chromatin in the hybrid cell achieved?
Our initial hypotheses were: (1) the identification of NB and GMC chromatin in the hybrid cell based on the differences in kinetochore composition or size. Asymmetric kinetochore composition was reported earlier, for example, sister chromatids in Drosophila male germline stem cells contain asymmetric levels of the centromere specific H3 variant (Centromere identifier (Cid) in flies)7,8; (2) Nuclear envelopes separate NB and GMC chromatin in the hybrid cell as it has been previously shown that fly neuroblasts undergo semi-closed mitosis9,10.
Although we have not observed any differences in the Cid between NB and GMC chromatin, we identified that NB Cid is connected to the active interphase Microtubule Organizing Center (MTOC), located close to the apical NB cortex. This apical MTOC-Cid connection keeps the NB chromatin from floating around the nucleus and prevents it from getting near the GMC chromatin, ensuring separation at least until metaphase. In addition to this, we found that NB-GMC hybrid cells contain independent nuclear envelopes, differing in size, which appeared to merge during late metaphase or early anaphase.
Based on our results, we propose that the separation, independent alignment, and segregation of NB and GMC chromosomes depends on (1) biased interphase centrosome activity in the NB, connecting the active apical MTOC with NB-chromatin throughout interphase and early mitosis, and (2) nuclear envelopes, imposing physical boundaries between the NB and GMC chromatin.
Where do we go from here?
The consequences of abrogated chromosome separation in either hybrid cells or zygotes should be studied through acute microtubule and nuclear envelope manipulations. We also plan to study the mechanisms and significance of the interphase microtubule-chromatin connections in neuroblasts.

References
- Ogle, B. M., Cascalho, M. & Platt, J. L. Biological implications of cell fusion. Nat. Rev. Mol. Cell Biol. 6, 567–575 (2005).
- Platt, J. L. & Cascalho, M. Cell Fusion in Malignancy: A Cause or Consequence? A Provocateur or Cure? Cells 8, 587 (2019).
- Reichmann, J. et al. Dual-spindle formation in zygotes keeps parental genomes apart in early mammalian embryos. 6 (2018).
- Kawamura, N. Fertilization and the first cleavage mitosis in insects. Dev. Growth Differ. 43, 343–349 (2001).
- Rieder, C. L. et al. Mitosis in vertebrate somatic cells with two spindles: Implications for the metaphase/anaphase transition checkpoint and cleavage. Proc. Natl. Acad. Sci. 94, 5107–5112 (1997).
- Heasley, L. R., Markus, S. M. & DeLuca, J. G. “Wait anaphase” signals are not confined to the mitotic spindle. Mol. Biol. Cell 28, 1186–1194 (2017).
- Ranjan, R., Snedeker, J. & Chen, X. Asymmetric Centromeres Differentially Coordinate with Mitotic Machinery to Ensure Biased Sister Chromatid Segregation in Germline Stem Cells. Cell Stem Cell 25, 666-681.e5 (2019).
- Dattoli, A. A. et al. Asymmetric assembly of centromeres epigenetically regulates stem cell fate. J. Cell Biol. 219, e201910084 (2020).
- Roubinet, C., White, I. J. & Baum, B. Asymmetric nuclear division in neural stem cells generates sibling nuclei that differ in size, envelope composition, and chromatin organization. Curr. Biol. 31, 3973-3983.e4 (2021).
- Katsani, K. R., Karess, R. E., Dostatni, N. & Doye, V. In Vivo Dynamics of Drosophila Nuclear Envelope Components. Mol. Biol. Cell 19, 3652–3666 (2008).
- Mohr, M., Zaenker, K.S., Dittmar, T. Fusion in Cancer: An Explanatory Model for Aneuploidy, Metastasis formation, and Drug Resistance. In: Pfannkuche, K. (eds) Cell Fusion. Methods in Molecular Biology, vol 1313. Humana Press, New York, NY (2015).
Follow the Topic
-
Communications Biology
An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.

Related Collections
With Collections, you can get published faster and increase your visibility.
Healthy Aging
Publishing Model: Open Access
Deadline: Dec 31, 2026
Cell death and inflammatory signalling
Publishing Model: Hybrid
Deadline: Oct 28, 2026

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in