Centrioles are frequently amplified in early B cell development but dispensable for humoral immunity
Published in Cell & Molecular Biology, Immunology, and Anatomy & Physiology
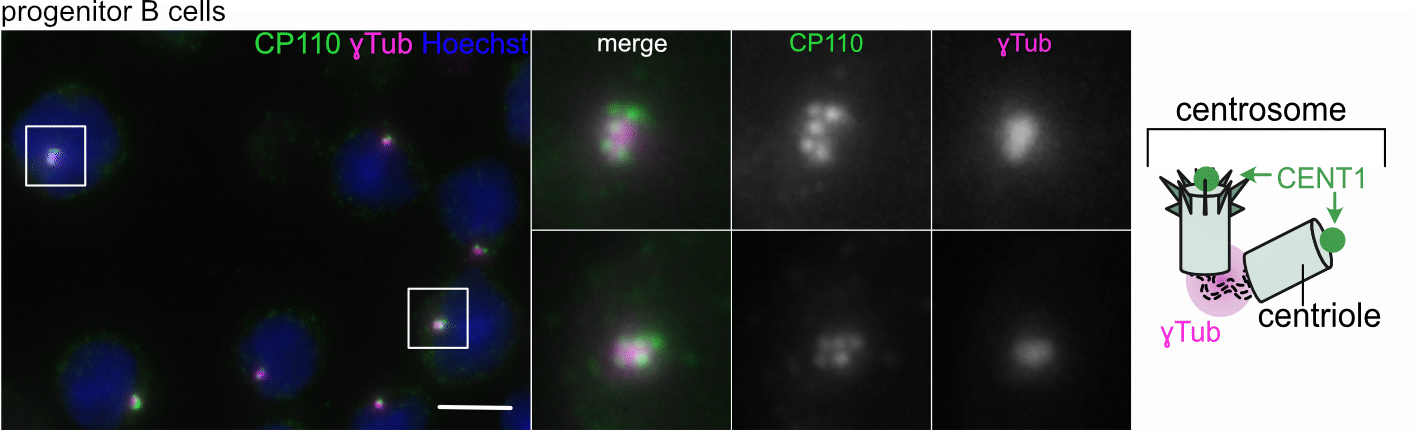
The current understanding
The centrosome is a highly conserved structure best known for its function in spindle assembly and chromosome segregation (Nigg and Holland, 2018). The two centrioles of each centrosome are usually duplicated only once per cell cycle, but centriole number aberrations are frequently observed in cancer and contribute to chromosomal instability (LoMastro and Holland, 2019). To prevent this, the proliferation of cells that have lost or amplified centrosomes is inhibited by the activation of the p53 tumor suppressor, either by the mitotic surveillance (Meitinger et al., 2016, Lambrus et al., 2016) or the PIDDosome pathway (Fava et al., 2017), respectively.
In recent years, our lab has focused on the role of the PIDDosome in cells that accumulate extra centrosomes naturally, such as polyploid hepatocytes (Sladky et al., 2020), or cardiomyocytes (Leone et al., 2024). However, extra centrosomes have also been observed in diploid cell types, including terminally differentiated dendritic cells, controlling innate immunity (Weier et al., 2022, Kiermaier et al., 2024). Therefore, we wondered, whether extra centrosomes are limited to terminally differentiated innate immune cells or if they are also found in adaptive immune system cells.
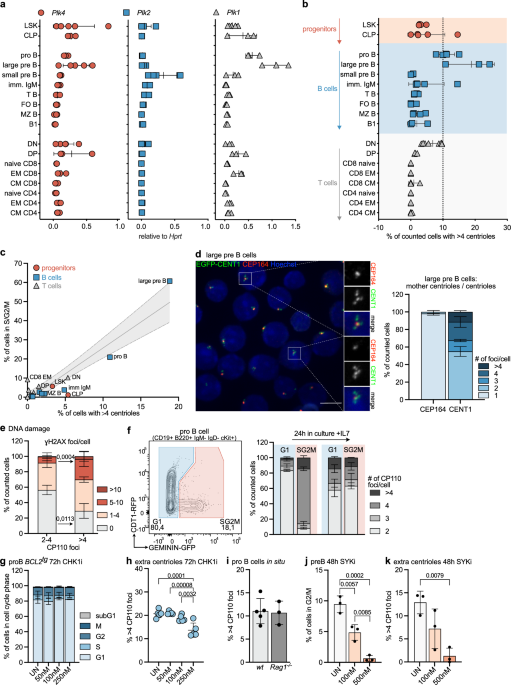
Centriole amplification in progenitor B cells is linked to proliferation and DNA damage (Fig. 1)
We found that centrioles were amplified in two types of proliferating progenitor B cells found in the bone marrow, pro and large pre B cells, with up to 25% of the latter displaying more than four centrioles. We linked centriole accumulation to delays in the G2/M transition, DNA damage, and premature centriole disengagement, regulated by CHK1 and PLK1 activity. Later stages of B cell development were no longer showing this phenomenon. We believe that centriole clearance in later stages is critical to maintain genome integrity, as early B cell malignancies often arise from pro and large pre B cells and commonly harbor centrosomal abnormalities. It is hence possible that early childhood malignancies could result from cells surviving with extra centrioles, fostering chromosome misseggregation and translocations, albeit this idea needs further testing. Moreover, despite our intense efforts, we do not know how centriole clearance is ultimately achieved during the course of B cell differentiation.

Figure 1. a. Quantification of IF images of cells with >4 CP110 foci. b. Model of centriole amplification. c. Quantification of ɣH2AX-foci in pro B with 2-4 or >4 centrioles by IF d. GFP-CENT1 pre B sorting scheme and cell cycle histograms. e. Quantification of IF images of pro B treated with CHK1i (PF47736) with >4 CP110-foci. f. Plk1 mRNA was assessed by qRT-PCR.
Centriole loss arrests B cell development but does not affect humoral immunity (Fig. 2)
Furthermore, we examined whether progenitor B cells, which tolerate extra centrioles, could also withstand centriole loss. Inhibiting centrosome duplication using centrinone, a drug blocking a key regulator of this process, PLK4, triggered BCL2-dependent, p53-regulated apoptosis in progenitor B cells. This sensitivity is far from universal, as centriole loss in mature germinal centre (GC) B cells did not affect survival or plasmablast differentiation, nor immunoglobulin production. To confirm these findings in vivo, we collaborated with the AJ Holland group at the Johns Hopkins University, and found that B cell development was blocked at the pro/pre-B stage in mice lacking Plk4 in the B cell lineage, but co-depletion of USP28, rescued this effect. These centriole-depleted mice also failed to produce OVA-specific IgG1 antibodies, which could also be restored by co-depletion of USP28, needed to activate p53-induced cell death downstream of centrosome loss. This suggests, that despite the role of the centrosome in antigen extraction, mature B cells can expand and respond to antigens without functional centrosomes, possibly due to low TRIM37 expression and suppression of p53 by BCL6 in germinal centre B cells.
Our work sheds new light on the role of centrioles in B cell development and function, with implications for the rise and treatment of different B cell malignancies.

Figure 2. a. Quantification of IF images of cells with <2 CP110-foci of centrinone-treated wt. b. Cell death (TOPRO3+) cells assessed by flow cytometry. c. Pro/pre-B fraction in the bone marrow (BM). d. Fraction of mature B cells. e. Serum anti-OVA IgG1 was assessed by ELISA 13 days post immunization. f. Centrinone-treated germinal center B (GCB) cells with <2 CP110-foci. g. Viable (TOPRO3-) cells assessed by flow cytometry. h. Trim37 mRNA expression was assessed by qRT-PCR analysis in pro B cells and follicular (FO) B cells. i. Scheme of the mitotic-surveillance-pathway.
References
Fava, L.L. et al. (2017) ‘The PIDDosome activates p53 in response to supernumerary centrosomes’, Genes and Development, 31(1). Available at: https://doi.org/10.1101/gad.289728.116.
Lambrus, B.G. et al. (2016) ‘A USP28-53BP1-p53-p21 signaling axis arrests growth after centrosome loss or prolonged mitosis’, Journal of Cell Biology, 214(2), pp. 143–153. Available at:https://doi.org/10.1083/jcb.201604054.
Kiermaier, E. et al. (2024) ‘Amplified centrosomes—more than just a threat’, EMBO Reports [Preprint]. EMBO Press. Available at: https://doi.org/10.1038/s44319-024-00260-0.
Leone M et al. (2024) ‘The PIDDosome controls cardiomyocyte polyploidization during postnatal heart development Running Title: The PIDDosome controls cardiomyocyte ploidy’, bioRxiv, p. 2024.08.27.609375. Available at: https://doi.org/10.1101/2024.08.27.609375.
LoMastro, G.M. and Holland, A.J. (2019) ‘The Emerging Link between Centrosome Aberrations and Metastasis’, Developmental Cell. Cell Press, pp. 325–331. Available at: https://doi.org/10.1016/j.devcel.2019.04.002.
Meitinger, F. et al. (2016) ‘53BP1 and USP28 mediate p53 activation and G1 arrest after centrosome loss or extended mitotic duration’, Journal of Cell Biology, 214(2), pp. 155–166. Available at: https://doi.org/10.1083/jcb.201604081.
Meitinger, F. et al. (2020) ‘TRIM37 controls cancer-specific vulnerability to PLK4 inhibition’, Nature, 585(7825). Available at: https://doi.org/10.1038/s41586-020-2710-1.
Nigg, E.A. and Holland, A.J. (2018) ‘Once and only once: Mechanisms of centriole duplication and their deregulation in diseases’, Nature Reviews Molecular Cell Biology. Available at: https://doi.org/10.1038/nrm.2017.127.
Sladky, V.C. et al. (2020) ‘PIDDosome‐induced p53‐dependent ploidy restriction facilitates hepatocarcinogenesis’, EMBO reports, 21(12). Available at: https://doi.org/10.15252/embr.202050893.
Weier, A.-K. et al. (2022) ‘Multiple centrosomes enhance migration and immune cell effector functions of mature dendritic cells’, Journal of Cell Biology, 221(12). Available at: https://doi.org/10.1083/jcb.202107134.
Figures were edited from the original publication.
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Biosensing
Publishing Model: Hybrid
Deadline: Sep 30, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in