CRISPR-Cas-based identification of a sialylated Human Milk Oligosaccharides utilization cluster in the infant gut commensal Bacteroides dorei
Published in Microbiology
Breastmilk is a fascinating biological product, an ancient recipe shaped by powerful evolutionary forces for the optimal nourishment of the newborn. Synthetic milk formulas are a good alternative for infants that are not (exclusively) breastfed for any reason, but our attempts to mimic mother nature’s complexity and diversity are lacking. One seemingly unusual component of breastmilk are Human Milk Oligosaccharides, or HMOs in short, and as a group these diverse sugars are completely indigestible by the infant. This perplexing observation is mitigated by the fact that HMOs are actually broken down by the bacterial population residing in the infant gut, namely the infant gut microbiome. Thus, the third most abundant component in breastmilk which consists of 10-15 g/L does not nourish the infant itself, rather it supports the colonization and prosperity of specific bacterial genera in its gut. In the context of establishment of the infant gut microbiome in the first months-years of life and the acquisition of bacteria from the environment, HMOs are certainly interesting to explore.
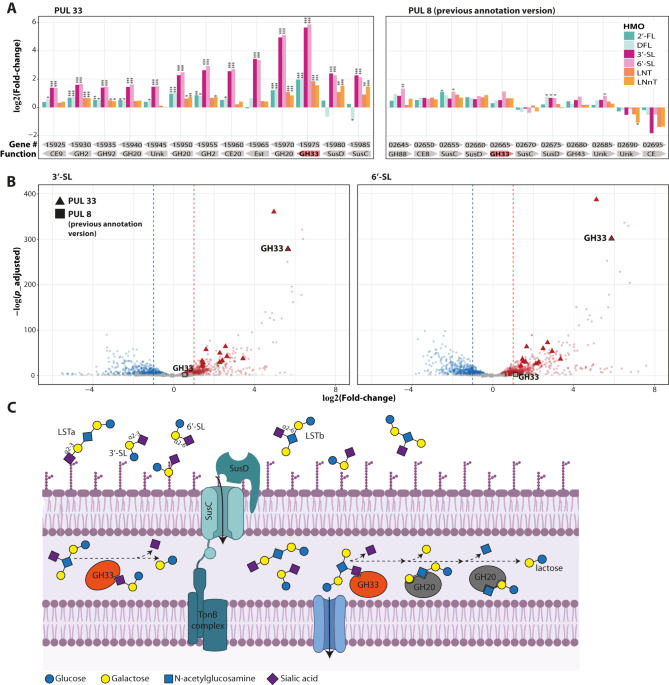
Our lab studies the development and establishment and developmen of the infant gut microbiome throughout the first months-years of life, and I chose to focus my research on Bacteroides, a bacterial genus that was somewhat overlooked in the context of HMO utilization although it is prevalent in the infant gut. Most research thus far has focused on Bifidobacterium species, the gold-standard HMO utilizers, but some computational evidence suggest that Bacteroides as well harbor genes that could break down the carbon bonds in HMOs, with additional studies showing experimentally that B. thetaiotaomicron and B. fragilis can grow on HMOs. However, mechanistic studies are lacking and the proposed pathways accounting for HMO utilization by those strains were suggested based on transcriptional analysis rather than genetic approaches. Similarly, using RNA sequencing, we found that a specific genomic cluster in a Bacteroides dorei isolate, polysaccharide utilization locus (PUL) 33, is entirely upregulated when growing on HMOs with a sialic acid terminal residues as the sole carbon source. While this strongly suggests that PUL33 has a central role in utilization of sialylated HMOs, we wanted more.
The genetic manipulation of my favorite B. dorei isolate turned out to be more technically complicated than the manipulation of commonly used lab strains I came across along the road, which was intimidating as a young PhD student in the lab. Bacteroides in general are difficult to genetically modify as they are anaerobes, usually slow-growing, and the genetic machinery constructed for more classic E. coli genetic engineering tends to be incompatible in Bacteroides. In addition, working with natural isolates (from breastfeeding infant stool samples) rather than well-characterized type-strains hinders genetic manipulation attempts even further, as allelic-exchange based methods are usually just not efficient enough, and the construction of a background mutated strain for each isolate is infeasible. Hence, we turned to CRISPR-Cas based genetic engineering for Bacteroides, published in 2022 (Zheng et al., ACS Synth Biol doi:10.1021/acssynbio.1c00543).
A while later, we managed to get the CRISPR-based genetic manipulation working for Bacteroides isolates in our lab, and knocked out the first four genes encoded in PUL33. The mutant strain, B. dorei Δ5k-PUL33, was unable to grow on sialylated HMOs (Fig 1) but growth on other HMO structures was preserved, indicating that PUL33 is required specifically for the break-down of HMOs containing a sialic acid. When complementing the four knocked-out genes back into the mutant strain, growth on sialylated HMOs was restored, establishing a genotype-phenotype link, but a cooler experiment showed that these four genes were enough to drive HMO utilization even in another Bacteroides strain, B. uniformis CL03T12C37, for which the WT strain cannot grow on sialylated HMOs (Fig 2).


Finally, using partial complementation essays, we showed that out of the four knocked-out genes, the gene encoding for the sialic acid-specific glycoside hydrolase (GH) 33 gene is the central one. Even the transporters of the PUL33 cluster were found to be dispensable, but B. dorei and B. uniformis lacking the GH33 gene could not utilize sialylated HMOs. Looking into the protein structure of the GH33 enzyme with AlphaFold, it was similar to a previously crystallized B. thetaiotaomicron GH33 sialidase. The B. thetaiotaomicron enzyme has a wide groove around the active site, shown to account for a wide substrate specificity of various sialylated sugar structures, and the B. dorei enzyme seems similar in this respect, with additional catalytic residues conserved as well.
Studying Bacteroides HMO-utilization in a mechanistic manner, using a gene-by-gene basic science approach, can serve as a base for the understanding of more complex dynamics in the ecosystem of the infant gut, with the ultimate goal of developing better synthetic milk formulas. I look forward to utilizing the CRISPR-based genomic editing system established in our lab for additional Bacteroides strains and gene clusters.
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Biosensing
Publishing Model: Hybrid
Deadline: Sep 30, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in