Double trouble: two Legionella pneumophila effectors modify the same histone residue
Published in Microbiology

Double trouble: two Legionella pneumophila effectors modify the same histone residue.
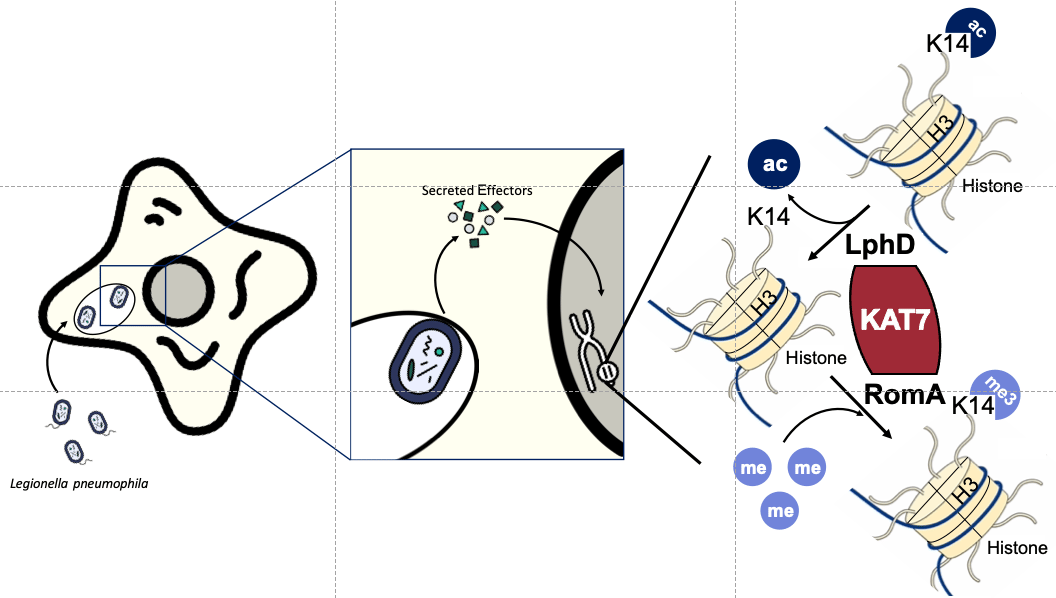
Legionella pneumophila is the causative agent of a severe form of pneumonia called Legionnaires' disease in humans. This bacterium has evolved several strategies to evade the immune system and survive within human cells1. To establish a replicative niche in this otherwise hostile environment, the bacterium acquired a large arsenal of secreted effectors that mimic eukaryotic proteins in the host cell during its evolution 2. By using different sets of effectors, depending on the colonized host or on the stage of its replicative cycle, L. pneumophila can manipulate diverse cell signaling pathways, mostly to delay cell response. One of its strategies involves manipulating the host epigenetic landscape to favor bacterial survival. Indeed, L. pneumophila secretes at least four nucleomodulins - bacterial effectors targeting the nucleus3. One of them is a eukaryotic-like SET-domain methyltransferase, called RomA, that specifically targets and methylates lysine 14 on histone H3 (H3K14), leading to a decrease in gene expression of several immune response genes4. However, in eukaryotic cells H3K14 is usually known to be acetylated, so the question that comes up is, “Are the bacteria actively involved in the process of deacetylation?”.
To address this question, we first searched the genome of L. pneumophila strain Paris and identified a perfect candidate gene, lpp2163 that encodes a 424 amino acid long protein carrying a predicted histone-deacetylase (HDAC)-domain. HDACs are enzymes that catalyze the removal of functional acetyl groups from the lysine residues of both histone and non-histone proteins5. We thus named this protein LphD (Legionella pneumophila histone Deacetylase) and started its enzymatic characterization. We show that LphD deacetylates lysine residues and that this activity can be inhibited by Trichostatin A (TSA), a broad range HDAC inhibitor. To gain insight into the protein structure we worked together with Mathilde Ben Assaya and Annemarie Wehenkel (Institut Pasteur). Using Alphafold they have shown that LphD shares high levels of similarity with eukaryotic HDACs, including several catalytic residues and an active site tyrosine. Moreover, they show that structurally, the most likely histone target was H3K14 since no other tested lysine residue fits as well as H3K14 into the binding pocket of LphD. In collaboration with Jérémy Berthelet and Fernando Rodrigues-Lima (Université de Paris) we confirmed, by quantifying deacetylation of histone H3 peptides by HPL C a preference of LphD for H3K14. This activity was conformed on short histone peptides, and on complete histone octamers.
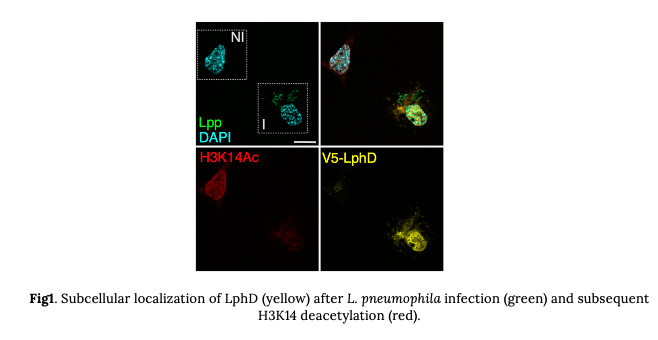
These results suggested that we were on the right track. We thus started looking at the role of LphD during infection. We first showed that LphD is a secreted via the L. pneumophila type IV secretion system (T4SS) Dot/Icm and that it localizes to the host cell nucleus. However, the exact mechanism of this nuclear localization is not known. On a side note, nuclear localization is not a given characteristic of histone deacetylases. In contradiction to their name, several histone deacetylases do not necessarily target the nucleus (or histones), some of them have well described non-histone targets. However, LphD is localizing to the nucleus during infection even though the nuclear localization signal was not identified. In the nucleus LphD decreases H3K14ac levels during infection with wildtype bacteria (Fig.1), but not a lphD knockout strain.
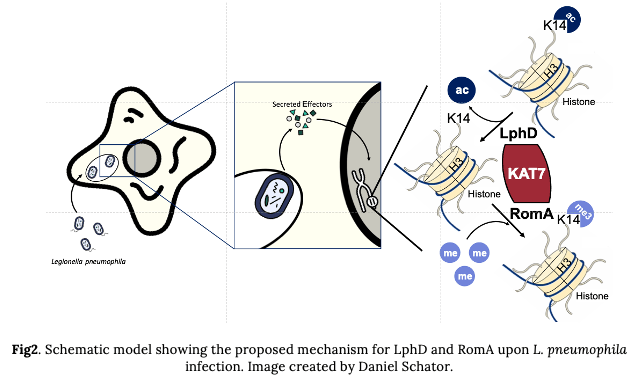
In addition, intriguingly, we also saw a drastic decrease in H3K14me (the mark regulated by RomA) when LphD was missing. This was the first sign that these two effectors could regulate the modification of H3K14 in synergy. The second key observation we made was that LphD and RomA interact with each other, pointing to a putative partnership in binding host chromatin. Such a complex formation at the chromatin level was confirmed by co-IP and ChIP experiments. We then tried to find the interaction partners of LphD in the host cell, that could drive its chromatin targeting. By performing GFP-pulldown from transfected cells (EGFP vs. EGFP-LphD) followed my mass spectrometry, we identified components belonging to two different eukaryotic HDAC-complexes (NuRD and Sin3) and several other epigenetic regulators. One of them, the histone acetyltransferase KAT7 caught our interest. In fact, KAT7 is the catalytic subunit of the HBO1 complex and is known as one of the main regulators of H3K14ac. So, we decided to focus on Kat7 and could show that LphD does indeed interact with several components of the HBO1 complex, including KAT7, in a context of L. pneumophila infection. In addition, RomA is also able to bind KAT7, further underlining the cooperation of LphD and RomA (Fig.2).
For sure at this point, you are wondering about the influence of LphD on bacterial replication within host cells. It was previously shown that a ∆romA strain has a deficit in intracellular replication in human cells (monocytic THP- 1), as well in protozoa (Acanthamoeba castellanii – a natural host of L. pneumophila). We performed the same assays using a ∆lphD strain which led to similar results: the knockout strain had a small but consistent deficit in intracellular replication. However, what happened next was probably the most unexpected and mind-boggling result of this project. When we looked at the behavior of the double mutant (∆lphD ∆romA) we expected a similar phenotype to the two single knockouts or maybe even a slight cumulative effect, but what we saw was that the double knockout partially reverted to the wildtype phenotype. This meant that for the bacteria it seemed to be more preferable to have neither of the two proteins than only one of them. This result was so consistent that we could even complement this phenotype. When reintroducing one of the two proteins into the double knockout, the bacteria showed again a decrease in intracellular replication and this decrease was dependent on the catalytic activity of the proteins, since complementation with catalytically dead versions did not lead to this decrease. The exact mechanism of this interaction is still an enigma, but a very exciting one to say the least. This contrast also led to the term para-effectors, from the Greek παρα (para), meaning besides but also contrary to, highlighting the ambivalence of these two effectors and their interdependence.
References
- Mondino, S. et al. Legionnaires’ Disease: State of the Art Knowledge of Pathogenesis Mechanisms of Legionella. Annu Rev Pathol 15, 439–466 (2020).
- Gomez-Valero, L. et al. More than 18,000 effectors in the Legionella genus genome provide multiple, independent combinations for replication in human cells. Proceedings of the National Academy of Sciences 116, 2265–2273 (2019).
- Bierne, H. & Pourpre, R. Bacterial Factors Targeting the Nucleus: The Growing Family of Nucleomodulins. Toxins (Basel) 12, 220–19 (2020).
- Rolando, M. et al. Legionella pneumophila effector RomA uniquely modifies host chromatin to repress gene expression and promote intracellular bacterial replication. Cell Host Microbe 13, 395–405 (2013).
- Seto, E. & Yoshida, M. Erasers of histone acetylation: the histone deacetylase enzymes. Cold Spring Harb Perspect Biol 6, a018713–a018713 (2014).
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in