Effects of presenilin-1 familial Alzheimer’s disease mutations on γ-secretase activation for cleavage of amyloid precursor protein
Published in Neuroscience

A Brief Introduction
γ-Secretase is an intramembrane aspartyl protease complex composed of four components Nicastrin (NCT), Aph-1, Pen-2, and Presenilin-1 (PS1). PS1 is the catalytic component of γ-secretase, “the proteasome of the membrane” which carries out intramembrane proteolysis of more than 150 peptide substrates, including amyloid precursor protein (APP), via two conserved aspartates, D257 and D385. Dominant missense mutations in PS1 can cause early-onset familial Alzheimer’s disease (FAD), a deadly chronic neurodegenerative disorder. Although disease-causing PS1 mutations were first identified over 25 years ago, exact pathogenic mechanisms of FAD mutations remain unclear.
Molecular dynamics (MD) has served as a powerful “Computational Microscope” for simulating atomic structural dynamics of biomolecules. However, conformational changes of the peptide substrate and γ-secretase during substrate cleavage of this well-known slow enzyme occur over long timescales (hours) and are beyond the reach of conventional MD simulations (typically microseconds). We have applied a novel Gaussian accelerated molecular dynamics (GaMD) method to address this challenge. GaMD has been shown to accelerate biomolecular simulations by orders of magnitude and thus called a “Time-Accelerated Computational Microscope”. Through GaMD enhanced simulations in previous studies, we have built dynamic models for activation of the wildtype (WT) γ-secretase and substrate cleavage of APP. First, the two PS1 catalytic aspartates are poised with ~7-8 Å distance between their Cγ atoms to recruit a water molecule for proteolysis through hydrogen bonds. Second, the carbonyl group at the cleavage site of APP form another hydrogen bond between the carbonyl oxygen and carboxylic side chain of protonated aspartate in PS1 for proteolysis.
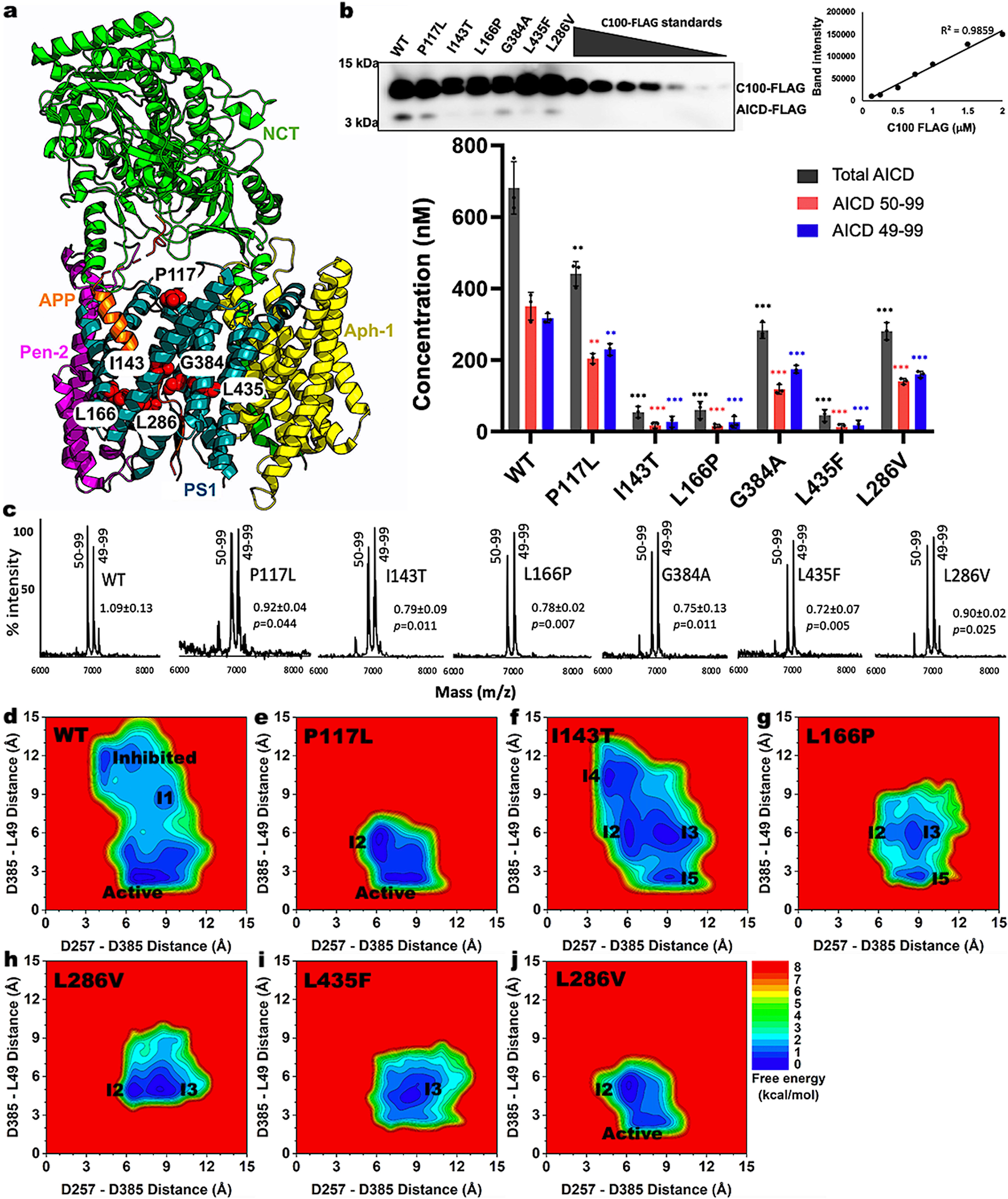
In our recent work published on Communications Biology, we have presented the first dynamic models for cleavage of APP by PS1 FAD mutants of γ-secretase, which were highly consistent with mass spectrometry (MS) and western blotting biochemical experiments. Through quantification of the total APP intracellular domain (AICD) species produced by WT and PS1 FAD mutant γ-secretase, our biochemical experiments revealed significantly reduced ecleavage of APP by the PS1 FAD mutants compared to the WT γ-secretase (Fig. 1b). GaMD simulations were carried out in parallel to explain the biochemical experimental data in atomistic detail. Different important low-energy conformational states were identified for each simulation system of γ-secretase from the free energy profiles calculated from GaMD simulations. This allowed us to deduce the effects of PS1 FAD mutations on the proteolytic activity of γ-secretase.
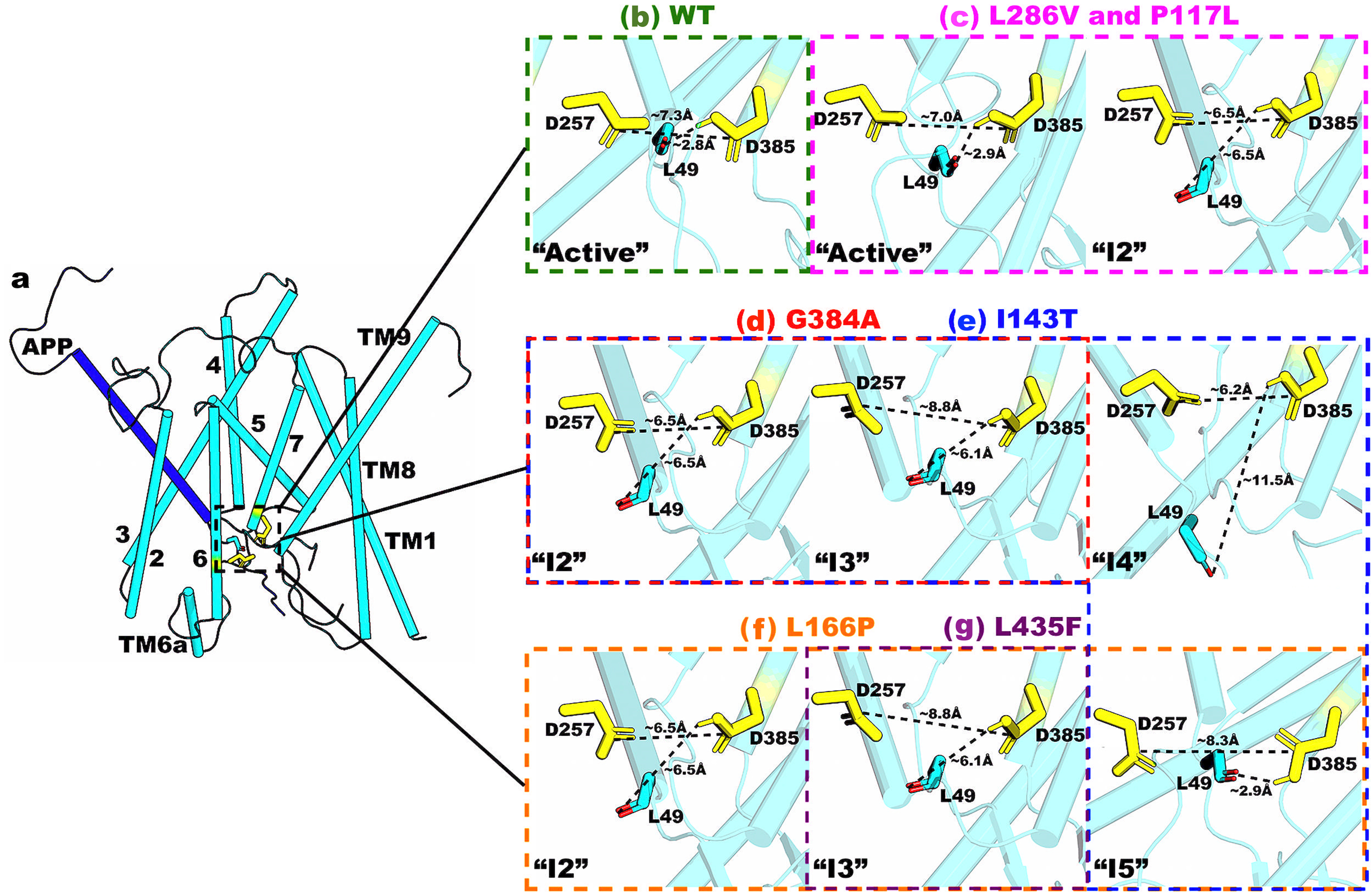
Flexibility of the active site played an important role in γ-secretase activation for ε cleavage of APP
We showed that flexibility of the active site played an important role in γ-secretase activation for ε cleavage of APP. In fact, the PS1 FAD mutants were found to stabilize the active site in PS1 and APP substrate. Even for PS1 FAD mutants such as P117L and L286V where the “Active” conformation was identified, the active site of PS1 exhibited reduced conformational space with respect to both the D257–D385 and D385–L49 distances relative to WT γ-secretase (Fig. 1d, 1e, and 1j). In addition, the P117L and L286V PS1 mutants sampled the intermediate 2 (or “I2”) conformation, in which the two catalytic aspartates moved too close to each other (Fig. 1e and 1j). Furthermore, the D257–D385 distance in the L286V PS1 FAD mutant complex could reduce further to ~4 Å. A ~4Å distance between D257–D385 signified a closed active site, in which a hydrogen bond was formed between the two catalytic aspartates and no water molecule could be recruited for γ-secretase activation.
PS1 FAD mutations hindered APP from proper orientation in the active site and/or disrupted the distance between the catalytic aspartates
The I143T PS1 FAD mutant sampled four important low-energy intermediate conformations, including “I2”-“I5” (Fig. 1f). The presence of the “I2” and “I4” conformations indicated that I143T was able to prevent APP from being properly located in the active site for its ε cleavage. On the other hand, this FAD mutant appeared to disrupt configuration of the catalytic aspartates given its adaptation of the “I3” and “I5” states. Therefore, the I143T PS1 FAD mutant could either prevent the APP substrate from aligning within the active site or disrupt the catalytic aspartate distances.
Three intermediate conformations were identified from the free energy profile of the L166P mutant, including “I2”, “I3”, and “I5” (Fig. 1g). As described above, the presence of “I2” and “I3” states suggested that the FAD mutation perturbed the APP substrate in the active site, while the presence of “I3” and “I5” states suggested that this mutation increased the D257–D385 distance. However, given the dominance of the “I5” and “I3” state compared to “I2”, the primary effect of the L166P FAD mutant appeared to be disrupting the D257 and D385 distance. The primary effect of the L435F PS1 FAD mutant was similar to that of the L166P as it mostly sampled the “I3” and “I5” states (Fig. 1i). This was to be expected as residue L435 in PS1 is located between the two catalytic aspartates D257 and D385. Its mutation to a bulkier residue such as phenylalanine created steric clashes within the PS1 active site, thereby increasing the D257–D385 distance.
Summary
In conclusion, the PS1 FAD mutations reduced the γ-secretase proteolytic activity by hindering APP residue L49 from proper orientation in the active site and/or disrupting the distance between the catalytic aspartates. Our findings (published on Communications Biology) provided important mechanistic insights into how PS1 FAD mutants affect structural dynamics and enzyme-substrate interactions of γ-secretase and APP.
Check out our related publication here: https://www.nature.com/articles/s42003-023-04539-1
Follow the Topic
-
Communications Biology
An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.

Your space to connect: The Psychedelics Hub
A new Communities’ space to connect, collaborate, and explore research on Psychotherapy, Clinical Psychology, and Neuroscience!
Continue reading announcementRelated Collections
With Collections, you can get published faster and increase your visibility.
Healthy Aging
Publishing Model: Open Access
Deadline: Dec 31, 2026
DNA repair and human disease
Publishing Model: Hybrid
Deadline: Oct 31, 2026



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in