Elucidating the metabolism of members of the Asgard archaea to help updating models on the origin of the eukaryotic cell
Published in Microbiology
Anja Spang & Thijs Ettema
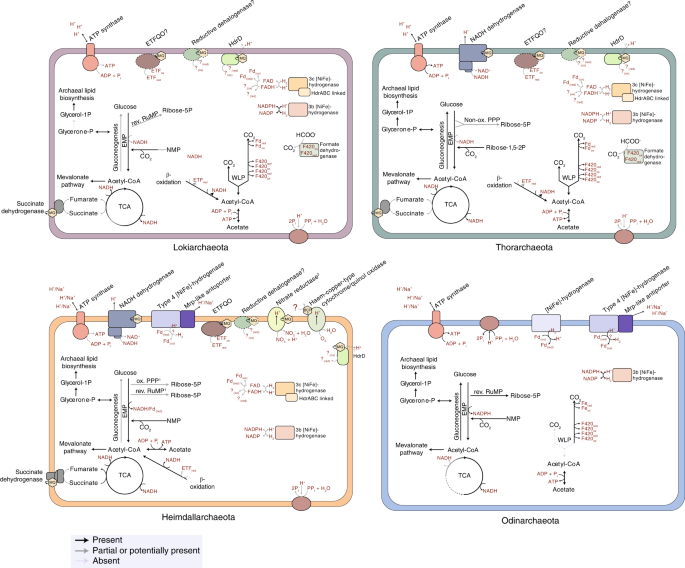
Only recently has the discovery of novel archaeal lineages enabled through the use of metagenomics, i.e. the sequencing of DNA directly isolated from environmental samples, helped to shed more light onto the elusive host of eukaryotic cells. In particular, such a sequencing approach has allowed the reconstruction of genomes of a so far uncultivated group of archaea, which was collectively referred to as the Asgard archaea and comprises lineages such as the Lokiarchaeota, Thorarchaeota, Heimdallarchaeota and Odinarchaeta (Spang et al., 2015; Zaremba-Niedzwiedzka et al., 2017). Organisms belonging to these lineages encode various proteins and protein domains that have previously only been found in eukaryotes but are generally absent from genomes of bacteria and other archaea. Furthermore, the placement of these lineages in the tree of life suggested that they represent the closest sister group of eukaryotes. Altogether, this research indicated, that the enigmatic host cell might have been an archaeon related to Asgard archaea.
But what was the driving force underlying the symbiosis between such an archaeal ancestor and the bacterial symbiont? Several previously formulated hypotheses, such as the hydrogen or symbiogenetic theory (Martin and Müller, 1998; Moreira and Lopez-Garcia, 1998) suggested that a metabolic interaction may have been central to eukaryogenesis. These scenarios are based upon the notion, that metabolic symbioses, i.e. syntrophic interactions between unrelated organisms, are common in nature and can lead to close associations based on the exchange of substrates or the direct transfer of electrons. For example, an interaction based on electron transfer characterizes various so called ANME consortia (McGlynn et al., 2015; Wegener et al., 2015), which consist of anaerobic methane oxidizing archaea and bacterial partners, the latter of which metabolize electrons generated by the archaea and thereby render growth on methane metabolically feasible.
However, given that previous scenarios were based on limited knowledge of the nature of the archaeal ancestor of eukaryotes, we decided to investigate the metabolic repertoire of extant members of the Asgard archaea and based on this knowledge formulate an updated model on the origin of the eukaryotic cell, which we refer to as the “Reverse flow model” (https://www.nature.com/articles/s41564-019-0406-9). In this model, we suggest that the archaeal ancestor of eukaryotes may have consumed organic substrates and generated reducing equivalents, that were metabolized syntrophically by the bacterial partner organism. The eukaryotic-like signature proteins of Asgard archaea, which among others are related to cytoskeleton and trafficking machinery proteins, may have played a role in establishing a tighter interaction between the archaeal host and bacterial partner(s). Over time, such a symbiotic interaction may have evolved into a more intricate relationship and ultimately into the loss of the integrity of the original partners.
While we think that this scenario seems best compatible with current knowledge on the genome repertoire of members of the Asgard archaea, the further characterization of this diverse archaeal lineage will be crucial to refine the steps that have led to the origin of the first eukaryotes more than 2 billion years ago. At the same time, the herein inferred metabolic features provide important information that can guide the targeted enrichment of Asgard archaea. Clearly, the ability to grow these archaea in a laboratory setting will be essential for characterizing the physiology and cell biology of these archaea and testing hypotheses on their ecology and interaction with other organism groups.
References
Lopez-Garcia, P. & Moreira, D. Open Questions on the Origin of Eukaryotes. Trends Ecol Evol 30, 697-708, doi:10.1016/j.tree.2015.09.005 (2015).
Lopez-Garcia, P., Eme, L. & Moreira, D. Symbiosis in eukaryotic evolution. J Theor Biol 434, 20-33, doi:10.1016/j.jtbi.2017.02.031
Martin, W. & Muller, M. The hydrogen hypothesis for the first eukaryote. Nature 392, 37-41, doi:10.1038/32096, (1998).
Martin, W. F., Garg, S. & Zimorski, V. Endosymbiotic theories for eukaryote origin. Philos Trans R Soc Lond B Biol Sci 370, 20140330-20140330 (2015).
McGlynn, S. E., Chadwick, G. L., Kempes, C. P. & Orphan, V. J. Single cell activity reveals direct electron transfer in methanotrophic consortia. Nature 526, 531-535, doi:10.1038/nature15512 (2015).
Moreira, D. & Lopez-Garcia, P. Symbiosis between methanogenic archaea and delta-proteobacteria as the origin of eukaryotes: the syntrophic hypothesis. J Mol Evol 47, 517-530 (1998).
Spang, A. et al. Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature 521, 173-+, doi:10.1038/nature14447 (2015).
Wegener, G., Krukenberg, V., Riedel, D., Tegetmeyer, H. E. & Boetius, A. Intercellular wiring enables electron transfer between methanotrophic archaea and bacteria. Nature 526, 587-590, doi:10.1038/nature15733 (2015).
Zaremba-Niedzwiedzka, K. et al. Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature 541, 353-358, doi:10.1038/nature21031 (2017).
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.





Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in