Epigenetic modulation via the C-terminal tail of H2A.Z
Published in Biomedical Research

Nucleosomal structure is, in general, repressive for transcription, replication and repair, thus one strategy of eukaryotic cells to regulate these activities involves de-repression by destabilizing or mobilizing particular nucleosomes. As such, the stability of nucleosomes is of utmost regulatory importance, and it is assumed to be modulated by posttranslational modifications (PTMs) on histones, by the reader proteins binding to them as well as by histone variant composition. In this context, destabilizing effects are expected in the case of activating functions, while nucleosome stabilization would suit repressive roles. In a puzzling manner, there are observations suggesting that the presence of the H2A.Z variant in nucleosomes can increase, or decrease nucleosome stability.
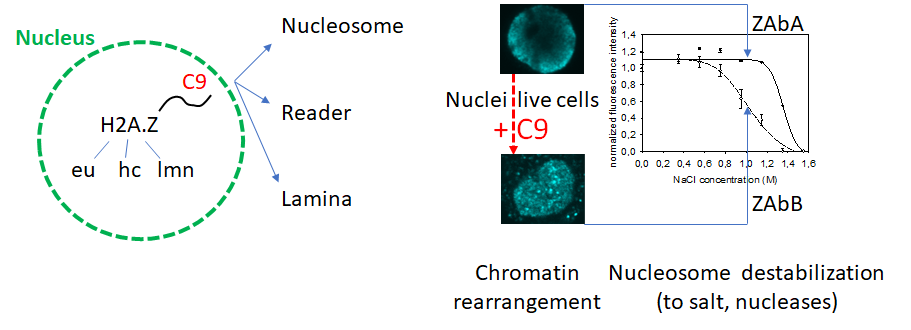
In this paper, nucleosome stability was measured in a robust in situ assay to assess its molecular determinants in H2A.Z-containing nucleosomes. The method (Imre L, et al. Nucleosome stability measured in situ by automated quantitative imaging. Scientific reports 7, 12734 (2017)) involves exposure of the nucleosomes in agarose-embedded nuclei to a concentration series of salt and measuring the histones of interest remaining in the nuclei by immunofluorescence and quantitative imaging, using antibodies specific for particular histones, histone variants or their posttranslational modifications (PTMs). This approach has led us to recognize a remarkable heterogeneity of H2A.Z-containing nucleosomes, independently from their isotype composition: three types can be observed, euchromatic (H2A.Zeu), heterochromatic (H2A.Zhc) and lamina-associated (H2A.Zlmn).
The primary observation was that a large fraction of H2A.Z detected by four different antibodies (H2A.Zhc) was released from the nucleosomes by salt together with H3, in contrast with the much less stably bound H2A.Zeu, H2A or H2B. Furthermore, this unusual behavior relied on the presence of the unstructured C-terminal chain of the histone variant, and was unaffected by H2AZ1 or H2A.Z2 isoform specificity or PTMs, as determined using cell lines expressing only particular forms of the variant and hydroxyapatite chromatography of reconstituted nucleosomes. In the absence of the tail, or upon addition of an excess of the tail peptide to the nuclei of control cells, the canonical H2A-like stability features prevailed or were readily restored as most of the H2A.Z-containing nucleosomes were observed as scattered foci in the nuclei rather than exhibiting the peripheral topography typical for H2A.Zhc. The organization of H3K9me3-marked constitutive heterochromatin as well as of the DNA itself within the native chromatin were also distinguishable in the nuclei of cells expressing wild type or tailless H2A.Z, and were rearranged upon addition of the tail peptide (C9) to isolated nuclei of cells harboring the wild type. Chromatin reorganization was accompanied by an increased overall sensitization to salt as well as to nucleases, i.e. destabilization. Increased MNase sensitivity could be demonstrated equally within H3K4me3-marked euchromatic and H3K9me3-marked heterochromatic domains. Thus, interactions involving a short H2A.Z peptide chain simultaneously determine the stability and accessibility features of chromatin involving the nucleosomes containing this histone variant, encompassing large chromatin regions in the nucleus.
When the tail wags the dog:
H2A.Z nucleosomes are heterogenous: they can be euchromatic (H2A.Zeu), heterochromatic (H2A.Zhc) or lamina associated H2A.Zlmn). Alternative binding of the H2A.Z C-terminal tail to the same or a neighboring nucleosome, to a reader protein recognizing the tail, or tethering chromatin to the nuclear lamina affect both nucleosome stability, as measured by in situ salt elution, and chromatin architecture. They both can be modulated by introducing the tail peptide (C9) to nuclei or live cells. There are two classes of commercial H2A.Z-specific antibodies: antibodies designated "ZAbA" recognize stable, heterochromatic, "ZAbB" binds to unstable nucleosomes of euchromatic localization.
When introduced into live cells C9 altered chromatin structure in a manner analogous to the result of a similar treatment of nuclei or the differences observed between cells expressing wild type and tailless H2A.Z, offering the possibility for targeted epigenetic manipulation. The observed changes and differences are best interpreted in terms of alternative binding of the tail to its molecular partners.
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Biosensing
Publishing Model: Hybrid
Deadline: Sep 30, 2026


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in