Find-and-replace CRISPR Genome Editing HDR2.0: A promising therapeutic Strategy
Published in Bioengineering & Biotechnology

Severe combined immunodeficiencies (SCIDs) are a set of devastating primary immunodeficiency disorders, attributed, in most part, to genetic mutations that disrupt T-cell development. SCIDs can also result in anomalies in B-cell and natural killer cell function and counts. If left untreated, SCID proves fatal within the first year of life. At present, the predominant method for definitively treating SCID patients is allogeneic hematopoietic stem cell transplantation (HSCT). Yet, the considerable challenges of locating a compatible donor and the potential immunological complications, such as graft-versus-host disease (GVHD), stand as significant hurdles in this treatment approach.
The advent of genome editing (GE) using targeted nucleases, particularly CRISPR-Cas9, has brought about a paradigm shift in gene therapy research, offering a potential remedy for such blood and immune system disorders1–4. The initial step in genome editing via the CRISPR-Cas9 system involves the creation of a site-specific double-strand break (DSB) by the Cas9 nuclease, which functions like a pair of molecular scissors. Subsequently, the cell’s own DNA repair machinery comes into play to repair this break. The repair process can follow either the non-homologous end joining (NHEJ) or homology-directed repair (HDR) pathways. In the NHEJ pathway, the repair process may result in the introduction of small insertions and deletions (INDELs). The intended outcome in this case is the targeted disruption, or knock-out (KO), of a specific gene. Conversely, in the HDR pathway, if a donor DNA is provided with short homologous arms and the precise sequence to be incorporated into the genome, the cell utilizes this donor DNA to repair the break via HDR. The desired result in this scenario is the precise correction of the targeted gene, effectively enabling GE to target nearly any gene in the genome. As a result, virtually every genomic disease becomes a potential target for therapeutic intervention5,6.
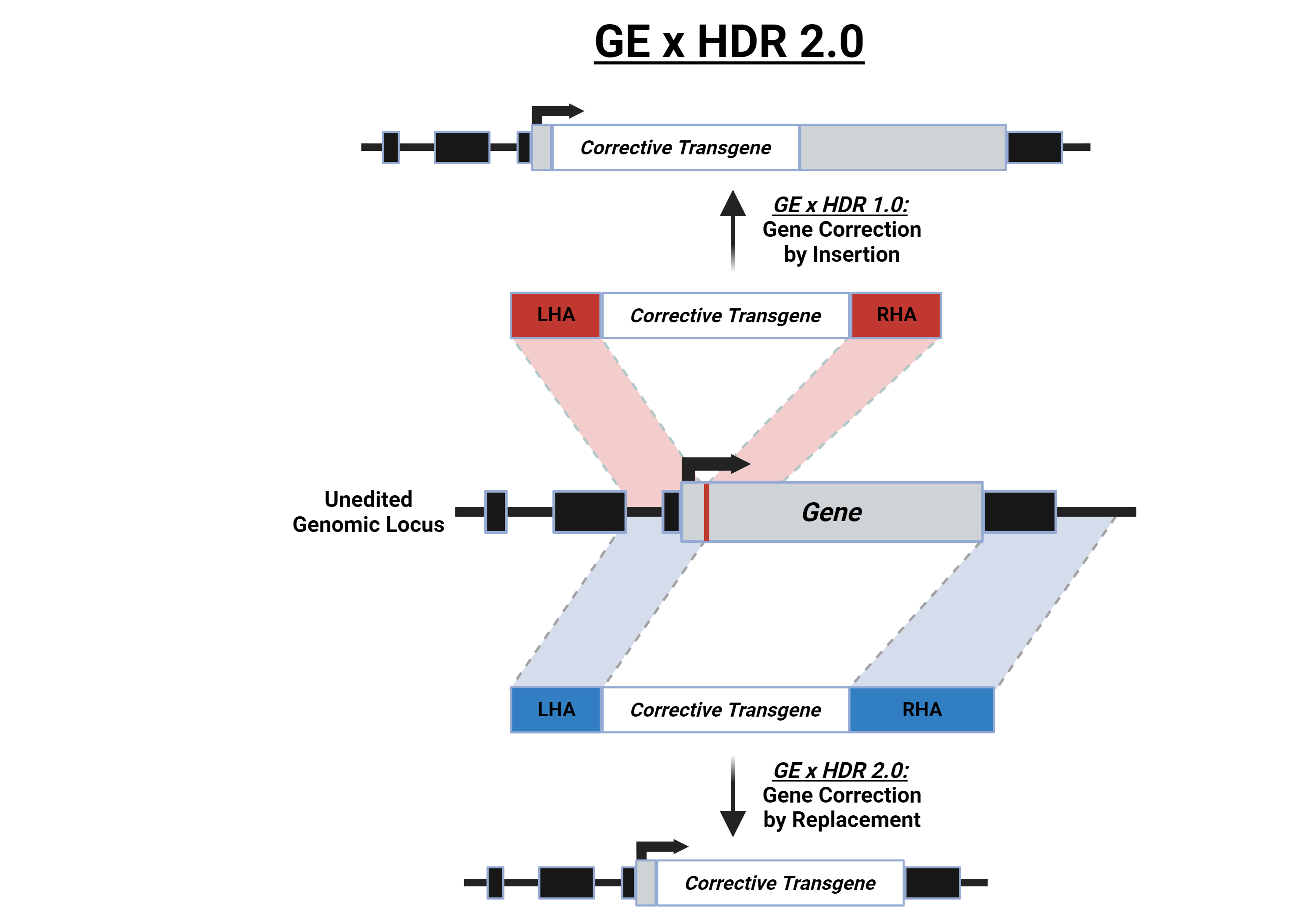
Among various genome-editing approaches, CRISPR-Cas9 HDR-mediated editing has emerged as a hopeful approach for site-specific incorporation of corrective transgenes to facilitate gene insertion. In specific subtypes of SCID, an alternative to hematopoietic stem cell transplantation (HSCT) involves viral vector-based gene therapy by gene addition strategies. However, in cases such as RAG2-SCID, this approach raises harmful risks and concerns. RAG2, along with RAG1, forms the nuclease responsible for cleaving DNA during V(D)J recombination. This process occurs during lymphocyte development, generating a highly diverse repertoire of antibodies and T-cell receptors. This process is orchestrated by a plethora of cis- and trans-acting regulatory elements in the RAG1/2 locus, which together create a 3D chromatin hub that acts as a super-enhancer that restricts the expression of the RAG genes, to the requisite developmental window in particular cell types. Potential risks following RAG viral-vector-based gene therapy approaches arise from possible ectopic expression of the RAG genes, which could lead to uncontrolled nuclease activity and the formation of harmful structural variations. For many genetic disorders, disease-causing mutations are spread across all coding exons and, therefore, it is difficult to develop a one-solution-fits-all strategy. Previous works to correct defunct genes have relied on inserting the corrective transgene into the nuclease-induced cut site (in the figure, top), however, this strategy does not enable for the complete conservation of endogenous regulatory elements both upstream and downstream to the cut site7. In this work, we propose a novel replacement strategy via CRISPR GE by HDR that we have termed GE x HDR 2.0: Find and Replace. We utilize CRISPR-Cas9-mediated GE in combination with recombinant adeno-associated serotype 6 (rAAV6) DNA donor vectors to precisely replace the endogenous RAG2 coding sequence (CDS) which is encoded by a single exon (in the figure, bottom). This approach enables us to correct all possible mutations in the CDS while preserving the native RAG2 expression regulatory elements both upstream and downstream to the replaced sequence. This strategy is particularly advantageous for the correction of tightly regulated genes like RAG2 and can be translated to the replacement of exons in genes with hot spot regions for disease-causing mutations since it is capable of conserving all endogenous regulatory elements and intronic sequences both 5’ and 3’ to the replaced fragment. In order to induce efficient HDR in a CDS replacement manner, it's essential to separate the distal homology arm from the cleavage site and align it with the sequence immediately downstream of the segment needing replacement. In this process, elongating the distal homology arm length in the donor is of paramount importance (note that in the figure below, the right homologous arm [RHA {blue}] for the replacement donor is longer than the RHA for the insertion donor [red], while the left homologous arm [LHA] is the same length for both donors). Thus, our innovative approach has the potential to replicate the natural expression levels of the endogenous gene with the corrective transgene’s expression levels and, therefore, prevent risks that are associated with unregulated expression.
In summary, in our study we present an innovative HDR-based gene-editing strategy, what we have called GE x HDR 2.0: Find and Replace. We developed a method to replace full CDSs or exons while maintaining endogenous and spatiotemporal regulatory elements or critical intronic sequences. We believe that this new generation of replacement gene-editing strategies can change the way the development of definitive therapies for disastrous and rare genetic diseases such as SCID have been approached to date.
https://www.nature.com/articles/s41467-023-42036-5
https://doi.org/10.1038/s41467-023-42036-5
References:
- Batzir, N. A., Tovin, A. & Hendel, A. Therapeutic Genome Editing and its Potential Enhancement through CRISPR Guide RNA and Cas9 Modifications. Pediatr Endocrinol Rev 14, 353–363 (2017).
- Shapiro, J., Tovin, A., Iancu, O., Allen, D. & Hendel, A. Chemical Modification of Guide RNAs for Improved CRISPR Activity in CD34+ Human Hematopoietic Stem and Progenitor Cells. Methods Mol Biol 2162, 37–48 (2021).
- Amit, I. et al. CRISPECTOR provides accurate estimation of genome editing translocation and off-target activity from comparative NGS data. Nat Commun 12, (2021).
- Allen, D. et al. High-Throughput Imaging of CRISPR- and Recombinant Adeno-Associated Virus-Induced DNA Damage Response in Human Hematopoietic Stem and Progenitor Cells. CRISPR J 5, 80–94 (2022).
- Allen, D., Rosenberg, M. & Hendel, A. Using Synthetically Engineered Guide RNAs to Enhance CRISPR Genome Editing Systems in Mammalian Cells. Front Genome Ed 2, 617910 (2020).
- Allen, D., Kalter, N., Rosenberg, M. & Hendel, A. Homology-Directed-Repair-Based Genome Editing in HSPCs for the Treatment of Inborn Errors of Immunity and Blood Disorders. Pharmaceutics 15, (2023).
- Iancu, O. et al. Multiplex HDR for disease and correction modeling of SCID by CRISPR genome editing in human HSPCs. Mol Ther Nucleic Acids 31, 105–121 (2023).
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in