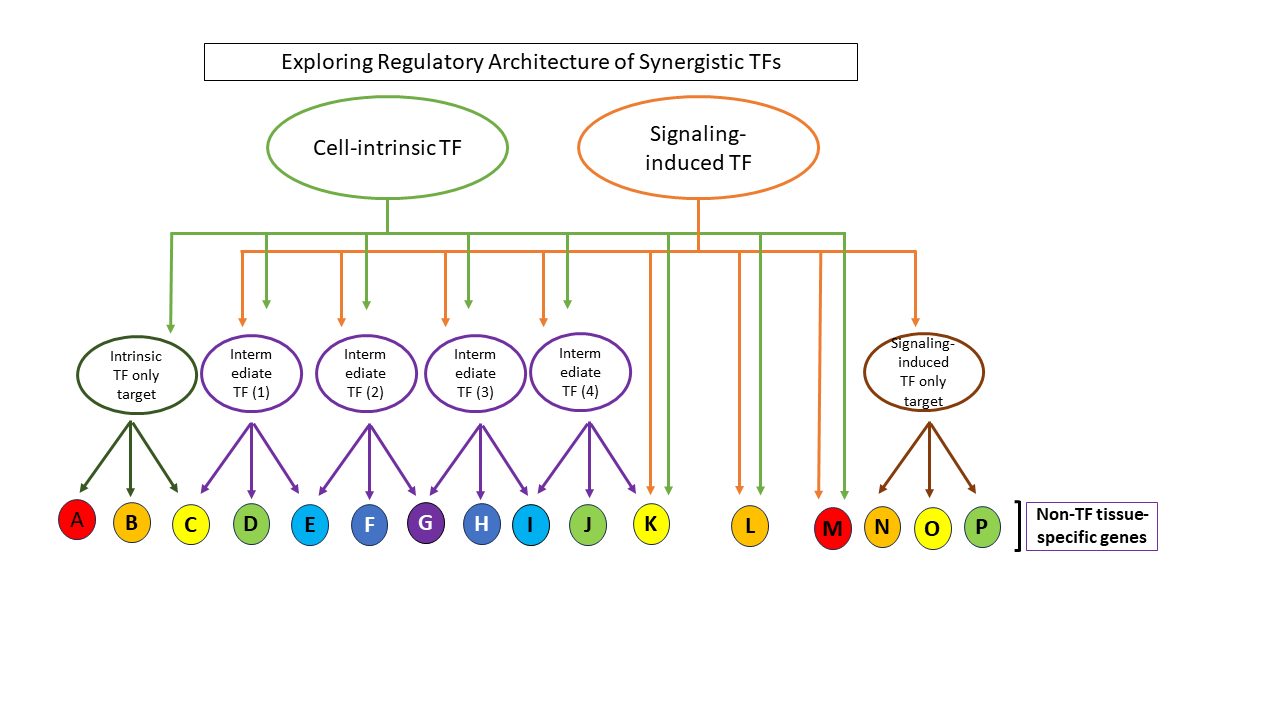
Transcriptional regulation remains one of the earliest major regulatory events in deciding whether a gene is expressed or not. Gene-specific transcription factors contribute greatly to the decision of whether a gene is activated or repressed. Changes in gene expression pattern can drive cell differentiation and cell fate changes in normal developmental processes and in disease. The availability of a cocktail of gene-specific transcription factors in a cell is a reflection of the lineage history of the cell, and sets the stage for temporal continuity of cell differentiation within a tissue. However, cell-cell signaling can activate a signaling cascade that will eventually induce a new set of gene-specific transcription factors (TFs) that could direct the cell to a new fate. Cell-cell signaling can also establish spatial patterns that contribute to the organization of a tissue. It is therefore interesting to know how existing TFs within the cell could integrate with TFs newly activated by cell-cell signaling to facilitate cell fate changes.
Drosophila melanogaster is a genetically tractable and fast-reproducing model organism to use for studying cell fate changes. Compared to mammalian systems, Drosophila in general has less redundancy in its key signaling components, eg. only one MEK gene (Dsor1) in the RAS signaling pathway. Flies also harbor functional homologs of 65% of human pathology-relevant genes. With the availability of next-generation sequencing methods, we could probe how intrinsic TFs integrate with extrinsic TFs in cell fate changes in vivo using transgenic flies. In our work titled “Synergistic activation by Glass and Pointed promotes neuronal identity in the Drosophila eye disc”, we discovered that the extrinsic ETS TF Pointed (Pnt), which acts downstream of Epidermal growth factor receptor (EGFR) signaling, synergistically activates neuronal genes in the developing eye with the intrinsic zinc finger TF Glass (Gl). We found activation of photoreceptor differentiation genes in a naïve environment, the wing disc, when we both overexpressed Gl and activated Ras signaling. This activation is lost in pnt mutant wing discs, indicating that the ability of Ras signaling to activate a neuronal differentiation program together with Gl requires pnt. To further understand the regulatory architecture in the developing eye, we performed Dam-ID Seq (a way to probe protein-DNA interactions) in in specific cell types in third instar larval eye discs. We found that Pnt and Glass co-bind to many targets in the genome of developing photoreceptors (PRs). These targets are enriched in predicted Gl and Pnt binding motifs. Pnt and Gl target genes include both intermediate TFs that contribute to PR development, as well as non-TF genes that build PRs. Existing research in the fly field also helped us to define an intermediate layer of TFs downstream of Gl and Pnt and upstream of PR structural genes. To further guide the construction of the regulatory architecture, we also used and performed additional RNA-Seq experiments to verify that the targets found in Dam-ID had differential gene expression upon perturbation of either Gl availability (glmutant) or EGFR signaling (Egfr temperature sensitive mutant). We propose that Gl and Pnt synergistically activate the PR program through a multi-layered network (A sample architecture is shown here, and our model is shown in fig. 7 in the paper).
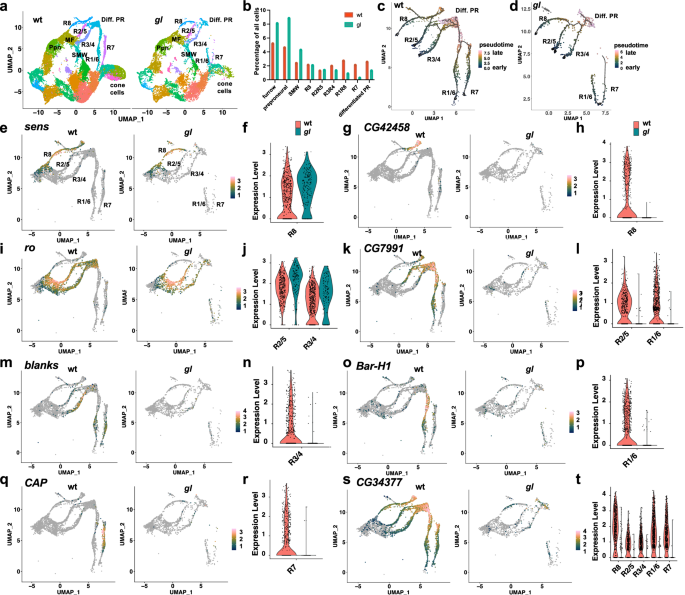
The defective eye of gl mutant flies was found over one hundred years ago, but using the recently developed single-cell RNASeq method, we were able to build a developmental trajectory of developing PRs in the white prepupal stage, ranging from undifferentiated to approximately 24 hrs after the induction of PR differentiation in the posterior of the eye disc. In this work, we published the single cell transcriptome of gl mutant eye disc cells at the white prepupal stage, capturing the earliest abnormalities in this mutant. With monocle3 trajectory analysis, we saw that the trajectories of the late-born PR subtypes R1, R6, and R7 are especially incomplete in gl mutants. Using Seurat integration and differential expression analysis in wild type and glmutant sc-RNA-Seq data, we found more PR-specific genes that are specifically perturbed in gl mutants. This dataset provides an example of photoreceptor subtype-specific changes in a TF mutant. We also included a github page to document our downstream scRNA-Seq analysis (DOI: 10.5281/zenodo.12770071).
Our work demonstrates that regulatory genes not only perform their designated function to regulate other genes, they also collaborate with their “colleagues” to enhance their own output in a cell differentiation setting. This is understandable as cell differentiation needs to be robust enough to push cells over epigenetic barriers for complete fate changes without set-backs. The two TFs we found to synergize came from two different sources; one is inherited as a general eye identity TF specific to the fly eye (Glass), and the other is a downstream effector of EGFR signaling (Pointed). The synergy between the two TFs integrates both the differentiation history of the cell and its current signaling status in space and time, and propels the cell to its future fate. This shows how evolution can generate a complex multicellular organism from a single cell with only limited numbers of signaling pathways and TFs. This synergy demonstrates that the combinatorial effects of TFs could yield extensive complexity without overloading our genome. It also reminds us that our genetic regulatory networks operate at precise spatiotemporal windows and dosages. The same EGFR signaling that produces the fly eye could turn it into a tumor if the signaling is not properly terminated. We observed that pnt gene transcription is attenuated late in the PR differentiation trajectory at the white prepupal stage, suggesting that this synergistic regulation does not persist when cell fate has been determined. Understanding how transcriptional regulation is regulated by both extrinsic signaling and intrinsic TFs is the first step towards developing combinatorial therapeutics for developmental disorders or other gene-misregulation-based pathologies.
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Biosensing
Publishing Model: Hybrid
Deadline: Sep 30, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in