Overcoming activity limitations of oligonucleotide catalysts through synthetic genetics
Published in Chemistry

The capacity of single-stranded nucleic acids to form not just duplexes with a complementary strand, but to fold into elaborate tertiary structures underpins the combinatorial power of nucleic acids: from deceptively simple chemical building blocks emerge complex structures with sophisticated biochemical functions.
Among these, RNA molecules with catalytic activity are not only at the heart of fundamental processes in modern biology - including protein synthesis and mRNA and tRNA processing - but are widely believed to have been instrumental at the origin of life. The discovery of such RNA enzymes (ribozymes), honoured with the award of the 1989 Nobel prize1, provided the first impetus for a deeper examination of the catalytic potential of natural nucleic acids as well as synthetic analogues.
Small trans-acting oligonucleotide catalysts capable of specific RNA cleavage – such as the hammerhead ribozyme, and laboratory-evolved analogous DNAzymes 10-23 and 8-172,3 – in particular have been intensively studied by synthetic chemical biologists for decades because of their great potential for medical and biotechnological applications. However, although some versions of these molecules are showing promise in diagnostic applications, the successful application of such catalysts as programmable, chemically-synthesised therapeutic agents for precision knockdown of disease-associated RNA transcripts has been hindered by the limitations inherent in natural nucleic acid chemistry. Although clinical trials have repeatedly demonstrated the safety and tolerability of ribozymes and DNAzymes in vivo, susceptibility to nuclease degradation, dependency on unphysiological concentration of metal ion cofactors, poor substrate accessibility within structured target RNAs and poor intracellular delivery have remained formidable obstacles to achieving clinical impact.
Systematic exploitation of insights gained from > 40 years of nucleic acid analogue chemistry has proven crucial for the translation of functional nucleic acid technologies to the clinic (antisense oligonucleotides (ASOs), silencing RNA (siRNA, RNAi) and synthetic mRNA)4. However, considerably less effort had gone into exploring modified nucleic acid chemistries in the context of ribozymes and DNAzymes with a view to addressing their limitations. Unequivocal demonstrations of specific cleavage activity, not just on short model substrates using high concentrations (≥10 mM) of divalent metal ions (typically Mg2+), but in particular on long, structured RNAs and full mRNA transcripts in physiologically relevant conditions (≤1 mM [Mg2+]) has remained elusive.
While in vivo (i.e. intracellular) RNA knockdown effects have been reported following DNAzyme transfections, such data must be interpreted with caution due to the number of pitfalls and sources of false-positives that exist when measuring such effects, as we outline in a recent Matters Arising article5. Indeed, effects in cells have been reported even when DNAzyme catalytic activity has been inactivated6 (as also observed with inactive ribozymes 30 years ago7), suggesting that they have the capacity to act as antisense oligonucleotides in vivo, reducing the level of target RNAs through simple base-pairing and recruitment of host silencing machinery (chiefly RNase H1) rather than intrinsic enzymatic RNA cutting.
The distinction between an ASO-like action and DNAzyme-mediated cleavage is crucial, as without it the principal advantages of DNAzyme technology - high cleavage specificity and host factor-independence - are lost.
Our interest in this area was sparked following our 2012 discovery that fully-modified artificial analogues of DNA and RNA not found in nature – known as xeno nucleic acids (XNA) – could not only serve as synthetic genetic polymers8 but could be evolved into functional structures (initially, XNA ligands or aptamers)9. We postulated that the chemical structure space of any information-carrying polymer capable of positioning functional groups in 3D could, in principle, support the evolution of enzymes and in 2015 we were able to confirm this through the directed evolution of a range of catalysts entirely composed of a variety of XNA chemistries, "XNAzymes", with RNA-endonuclease and RNA- and XNA-ligase activities10,11.
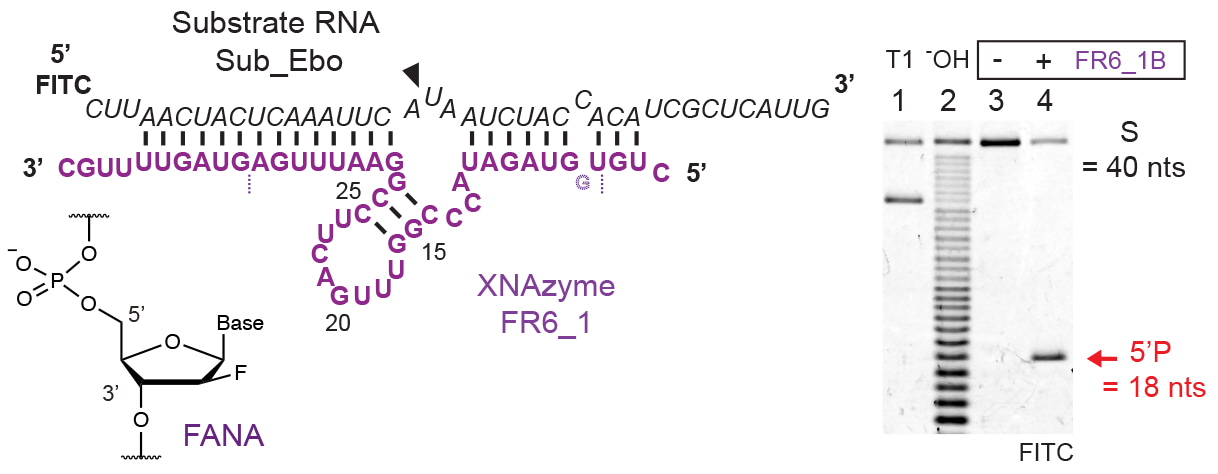
We reasoned that the capability of XNA chemistries such as 2’-deoxy-2’-fluoro-β-D-arabino nucleic acid (FANA) to form structures with enhanced stability compared with DNA or RNA might alleviate the requirement for divalent metal ions for folding and catalysis. Despite this, the first generation of RNA-cleaving XNAzymes exhibited poor activity on all-RNA substrates in physiological magnesium concentrations (0.5 - 1 mM Mg2+) nor could they be easily reprogrammed to cleave other target sequences; activity quickly dropped off as substrate binding arms were cumulatively mutated. However, after modifying our directed evolution workflow and selection strategies for the discovery of XNAzymes capable of cleaving more ‘realistic’ target RNAs under low [Mg2+], we were able to identify a new all-FANA catalytic motif - FR6_1 - that not only outperformed previous XNAzymes, but retained its RNA cleavage activity in physiological conditions. Furthermore, FR6_1 could be readily re-targeted to cleave a range of highly structured full-length (>5 kb) (m)RNA targets.
By exploiting the FR6_1 XNAzyme’s high RNA cleavage specificity, as well as the ‘programmability’ of substrate-binding arm sequences, we were able to demonstrate a key proof-of-concept of the potential power of the technology: cleaving realistic RNA transcripts or substrates with single-nucleotide precision, in this case cancer-associated mutants of classic oncogenes KRAS [G12D] (Fig. 1) and BRAF [V600E]. This enabled allele-specific mRNA knockdown of G12D (c.G35A) KRAS mRNA within hours of transfection into live cultured adenocarcinoma cells, both in a KRAS G12D homozygous and heterozygous genetic background, in the presence of wild-type (c.G35) mRNA.

A variant of the FR6_1 FANA XNAzyme targeted to codon 12 in the mRNA of the oncogene KRAS, a cancer-associated mutation hotspot. XNAzyme "FR6_1_KRas12B" specifically cleaves the full-length transcript encoding the common mutation c.35G>A [G12D], but not the wild type, in physiological conditions in vitro and inside live colon carcinoma cells. Measured by droplet digital RT-qPCR.
Although we find that FANA XNAzymes continue to induce a (less specific) antisense-mediated knockdown, we can now disentangle these contributions from (allele-specific) XNAzyme catalysis by development and validation of stringent controls, a catalytically-inactive point mutant of FR6_1 and an analogous variant of the classic 10-23 DNAzyme. This also involved substantial efforts to eliminate technical artefacts associated with commonly used strategies for cellular RNA workup and measurement of gene silencing by qRT-PCR that are not ideally suited for work with XNAzymes or XNA-modified DNAzymes. Due to their higher base-pairing stability and partial resistance to nuclease degradation these can remain active in cellular extracts during DNase treatment and subsequent RT steps, yielding false positive knockdown. Therefore, great care must be taken to remove them from mRNA preparations to ensure measurements truly reflect intracellular activity. Using these stringent controls, we demonstrate that approximately 50% of FR6_1 XNAzyme-mediated knockdown occurs due to the bona fide catalytic turnover of the XNAzyme inside cells, distinct from antisense-type effects.
Our results underline the potential of XNA technology to overcome some of the long-standing difficulties associated with oligonucleotide catalysts, in particular in enabling substantial functional gains under physiological conditions and against realistic targets, providing a foundation for the more systematic development of improved gene silencing agents based on FANA or other XNA chemistries. Indeed, chemistries even more robust than FANA, such as 2’OMe-, 2’-MOE-RNA or locked nucleic acids (LNA), with substantially reduced or abolished capacity to trigger RNase H, offer promise for the discovery of next-generation XNAzymes with ever more precise and long-lasting in vivo activities. Although technically challenging, incorporation of expanded nucleobases as well as backbone chemistry into XNAzyme selections also suggests a route to catalysts with a greater diversity of reaction mechanisms. Bringing mechanisms analogous to those employed by proteinaceous enzymes into the realm of oligo catalysts would provide substantial gains in catalytic turnover12 enabling improved knockdown and lower effective doses - as well as the exciting prospect of catalysts for reactions and chemical environments inaccessible to enzymes composed of natural polymers.
References:
1 The Nobel Prize in Chemistry 1989. https://www.nobelprize.org/prizes/chemistry/1989/press-release/
2 Faulhammer, D. & Famulok, M. Characterization and divalent metal-ion dependence of in vitro selected deoxyribozymes which cleave DNA/RNA chimeric oligonucleotides. Journal of Molecular Biology 269, 188-202 (1997). https://doi.org/10.1006/jmbi.1997.1036
3 Santoro, S. W. & Joyce, G. F. A general purpose RNA-cleaving DNA enzyme. Proceedings of the National Academy of Sciences of the United States of America 94, 4262-4266 (1997). https://doi.org/10.1073/pnas.94.9.4262
4 Gait, M. J. & Agrawal, S. Introduction and history of the chemistry of nucleic acids therapeutics. Methods in Molecular Biology 2434, 3-31 (2022). https://doi.org/10.1007/978-1-0716-2010-6_1
5 Taylor, A. I. & Holliger, P. On gene silencing by the X10-23 DNAzyme. Nature Chemistry (2022). https://doi.org/10.1038/s41557-022-00990-5
6 Young, D. D., Lively, M. O. & Dieters, A. Activation and deactivation of DNAzyme and antisense function with light for the photochemical regulation of gene expression in mammalian cells. Journal of the American Chemical Society 132, 6183-6193 (2010). https://doi.org/10.1021/ja100710j
7 Steinecke, P., Herget, T. & Schreier, P. H. Expression of a chimeric ribozyme gene results in endonucleolytic cleavage of target mRNA and a concomitant reduction of gene expression in vivo. The EMBO Journal 11, 1525-1530 (1992). https://doi.org/10.1002/j.1460-2075.1992.tb05197.x
8 Pinheiro, V. B., Taylor, A. I., Cozens, C., Abramov, M., Renders, M., Zhang, S., Chaput, J. C., Wengel, J., Peak-Chew, S.-Y., McLaughlin, S. H., Herdewijn, P. & Holliger, P. Synthetic genetic polymers capable of heredity and evolution. Science 336, 341-344 (2012). https://doi.org/10.1126/science.1217622
9 Taylor, A. I., Houlihan, G. & Holliger, P. Beyond DNA and RNA: The expanding toolbox of synthetic genetics. Cold Spring Harbor Perspectives in Biology 11, a032490 (2019). https://doi.org/10.1101/cshperspect.a032490
10 Taylor, A. I. & Holliger, P. Directed evolution of artificial enzymes (XNAzymes) from diverse repertoires of synthetic genetic polymers. Nature Protocols 10, 1625-1642 (2015). https://doi.org/10.1038/nprot.2015.104
11 Taylor, A. I., Pinheiro, V. B., Smola, M. J., Morgunov, A. S., Peak-Chew, S.-Y., Cozens, C., Weeks, K. M., Herdewijn, P. & Holliger, P. Catalysts from synthetic genetic polymers. Nature 518, 427-430 (2015). https://doi.org/10.1038/nature13982
12 Wang, Y., Liu, E., Lam, C. H. & Perrin, D. M. A densely modified M2+-independent DNAzyme that cleaves RNA efficiently with multiple catalytic turnover. Chemical Science 9, 1813-1821 (2018). https://doi.org/10.1039/C7SC04491G
Follow the Topic
-
Nature Chemistry
A monthly journal dedicated to publishing high-quality papers that describe the most significant and cutting-edge research in all areas of chemistry, reflecting the traditional core subjects of analytical, inorganic, organic and physical chemistry.




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in