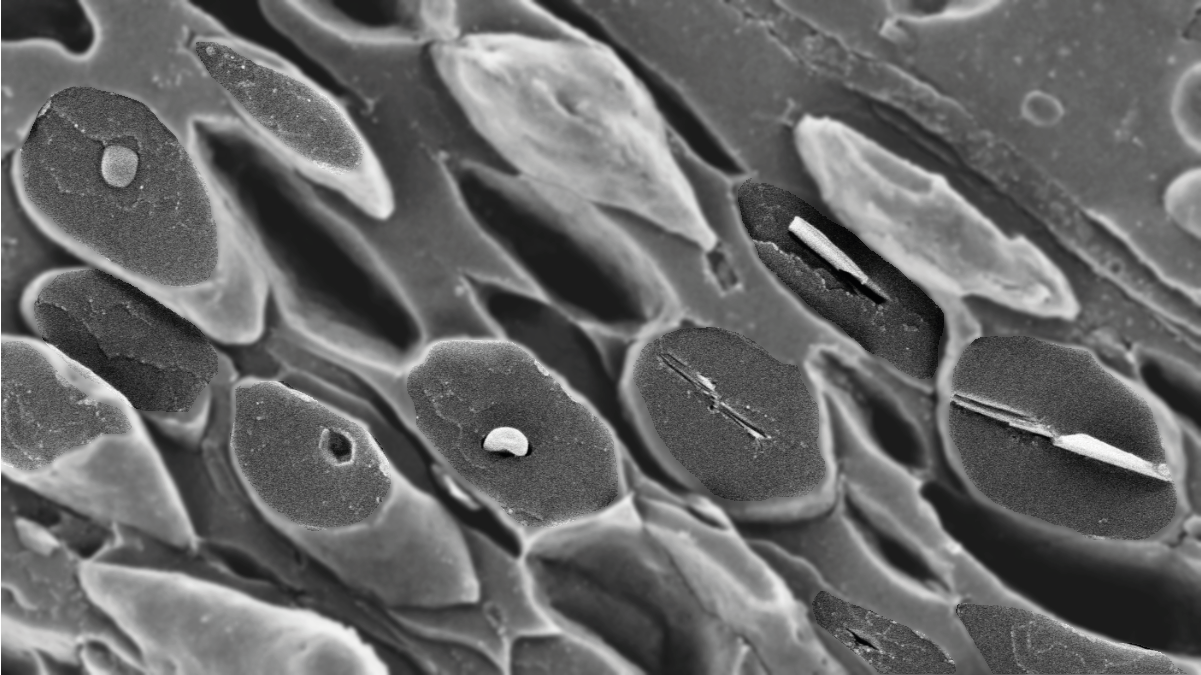

More than half a century ago, neuroscientist Michael Land peered through the eye of a scallop and saw an upside-down image of himself looking back. Land’s unnerving experience resulted in his discovery of a new type of eye – the concave mirrored eye1. Even today, most people don’t know scallops have eyes, let alone dozens to hundreds of them lining the tissue between the two valves. Land found that each one of these sub-millimeter eyes contains a concave mirror which focuses light, much like a reflecting telescope, onto the overlying retinae. The mirror is formed from alternate layers of high-refractive index guanine crystals and low-refractive index cytoplasm, creating a multilayer interference reflector2,3.
The unusual scallop eye was destined to become a star, not only of visual ecology, but of the field of organic biomineralization, which concerns the formation and function of biogenic molecular crystals. The electron microscopy techniques of Land’s days could not preserve the beam sensitive organic crystals, which instead appeared as angular voids in the tissue4. Over 50 years later, augmented by the advent of cryogenic scanning electron microscopy, which enables organic crystals to be imaged in their native state, Palmer and co-workers re-investigated this remarkable visual system. They unearthed previously unknown intricacies in the optical design of the scallops image-forming mirror which is constructed from a perfectly tiled mosaic of square guanine crystals5. The study concluded that understanding how organisms control the shape and arrangement of crystals could lead to the development of novel bioinspired optical devices.
One conundrum presented by the scallop is the symmetrical square shape of the crystals, a morphology formally prohibited by guanine’s monoclinic crystal symmetry. An electron diffraction study showed the square tiles are not single crystals but rather composites of three twinned domains, enabling the monoclinic guanine crystal to mimic higher symmetry structures6.
A second feature of the scallop’s guanine crystals, as well as those in fish, copepods and spiders, is the preferential expression of their highly reflective face - not manifest in crystals grown synthetically from water. In this paper, we delve deeper into how scallops regulate this morphology control. Our strategy was to follow guanine formation in developing juvenile scallops and to survey changes in crystal morphology during development. Guanine crystals form inside organelles called iridosome vesicles. We observed that early iridosomes, which do not yet contain a crystal, contain two pre-assembled fibrillar sheets. A crystal then nucleates in the space between these sheets and is guided by the sheets to grow in a specific direction that allows the crystal to express its high refractive index face. The sheets perform three essential functions related to crystal nucleation and growth: (i) they define a space in which crystal formation can occur within the iridosome, (ii) they provide an interface for guanine nucleation on the planar face of the molecule, and (iii) they cap the high refractive index faces of the growing crystals, preventing growth in the π-stacking direction. In addition, we determined that even prior to crystal nucleation the directionality of sheets in a juvenile scallop closely align with the curvature of the mirror in the adult scallop. In other words, crystal orientation is ‘pre-programmed’ into the early iridosomes.
The incredible crystal engineering capabilities of scallops are demonstrated by their ability to make thousands of identical, ultra-small square plate crystals (1.2 x 1.2 x 0.07 µm3) and arrange them into a highly ordered, functioning mirror that spans the entire diameter of their eye (200 to 1200 µm). Scallops achieve this feat by using pre-assembled macromolecular templates. The precise level of control over the size, shape, and arrangement of guanine crystals in scallop eyes are a testament to the effectiveness of macromolecular templates in molecular crystal growth. In our efforts to replicate this level of molecular crystal morphology control in the laboratory, we hope to harness this strategy and create novel molecular materials with desired properties.
The connection between freckles and scallops
The formation of iridosomes in scallops described in this study is strikingly similar to the well-documented morphogenesis of melanosomes in vertebrates. The common origin of pigment cells in vertebrates was first proposed by Bagnara and colleagues7 and the current leading hypothesis is that in all animals, pigment organelles originate from the endosomal system and are part of the lysosome-related organelle (LROs) family8. Until now, the ultrastructural features of iridosomes and their similarities to other LROs were not well understood. Our study helps to bridge this gap, by demonstrating that guanine, like melanin, forms on a template of pre-assembled fibrillar sheets that remains incorporated in the final organelle. By drawing parallels with melanin formation, we hope to gain new insights into the chemical composition of iridosomes, which is currently not well known.
References
- Land, M. F. Image formation by a concave reflector in the eye of the scallop, Pecten maximus. J. Physiol. 179, 138–153 (1965).
- Barber, V. C., Evans, E. M. & Land, M. F. The fine structure of the eye of the mollusc Pecten maximus. Zeitschrift fur Zellforsch. 76, 295–312 (1967).
- Land, M. F. A multilayer interference reflector in the eye of the scallop, Pecten maximus. J. Exp. Biol. 45, 433–447 (1966).
- Wagner, A. et al. Functional Molecular Crystals in Biology. Isr. J. Chem. 61, 668 – 678 (2021).
- Palmer, B. A. et al. The image-forming mirror in the eye of the scallop. Science. 358, 1172–1175 (2017).
- Hirsch, A. et al. Biologically controlled morphology and twinning in guanine crystals. Angew. Chemie - Int. Ed. 56, 9420–9424 (2017).
- Bagnara, J. T. et al. Common origin of pigment cells. 203, 410–415 (1979).
- Figon, F., Deravi, L. F. & Casas, J. Barriers and promises of the developing pigment organelle field. Integr. Comp. Biol. 61, 1481–1489 (2021).
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in