Teaching crop plants how to concentrate CO2
Published in Bioengineering & Biotechnology

Our lives rely on CO2 fixation, or conversion of CO2 into sugars and other carbon-based compounds. Every year, roughly 300 billion tons of atmospheric CO2 are fixed by photosynthetic organisms and made into useful things [1]. Look around — the world is full of carbon commodities produced by the process of CO2 fixation: clothing, plastics, fuels, and most importantly, food.
Over the last century, food supplies were able to match human population growth because of dramatic boosts in crop yields. Nevertheless, the recent drivers of improvements in yields, such as optimization of the partitioning of biomass into grains, have begun to plateau [2]. As the global population continues to grow, the rising demand for food is projected to outpace the increase in crop yields by 2050 [3]. Therefore, we need to develop new strategies to increase crop yields.
The major limiting factor for the growth of many crop plants is the rate at which they can uptake and fix CO2. In crop plants, as well as in almost every other photosynthetic organism, a key enzyme called Rubisco drives CO2 fixation, converting CO2 into organic compounds. Rubisco works most effectively at high concentrations of CO2, more than three times higher than the CO2 levels in our current atmosphere and in typical plant chloroplasts. Consequently, in many crops Rubisco does not achieve its maximum catalytic activity. This provides an attractive target for improving yields.
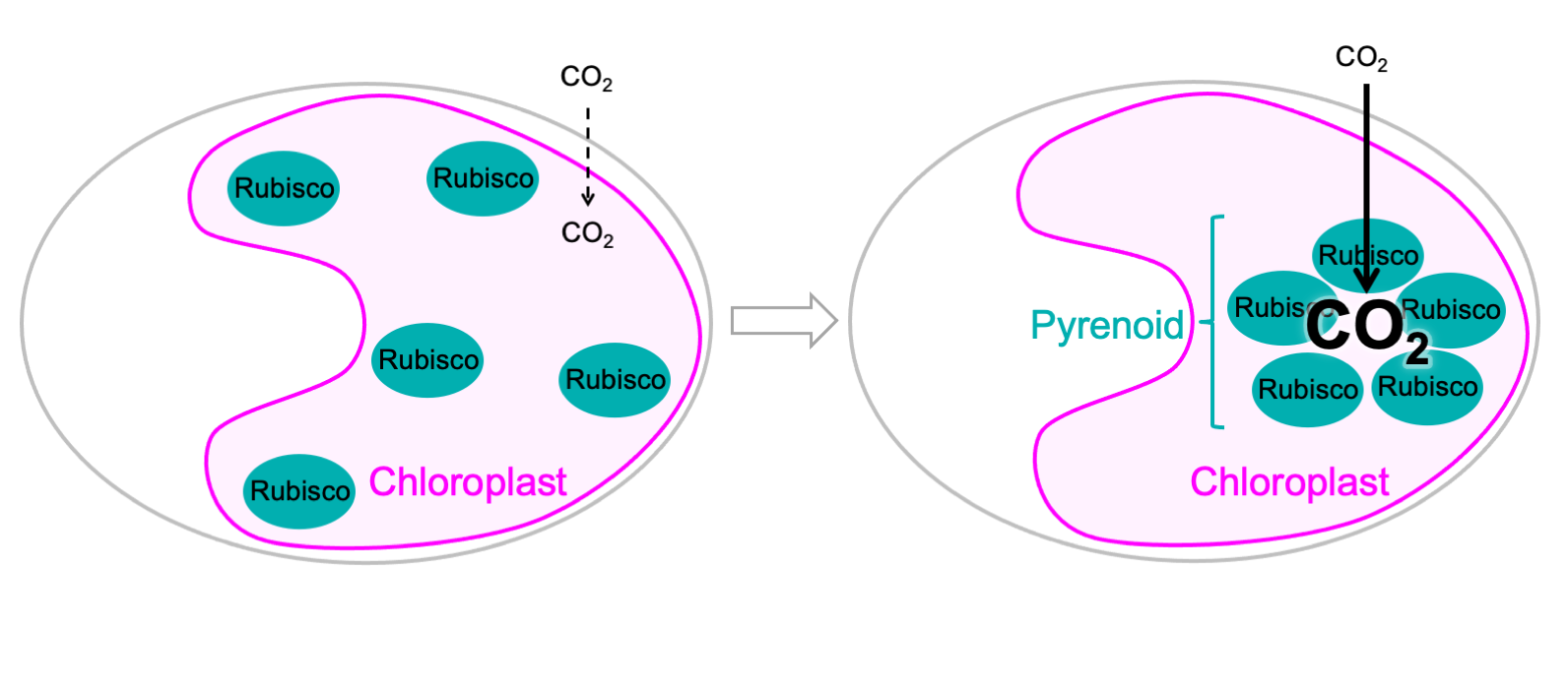
Can we supply Rubisco with locally concentrated CO2 to make it run faster? The answer is yes. In fact, many photosynthetic organisms have invented CO2-concentrating mechanisms (CCMs) to perform this trick [4,5,6]. A remarkable example can be found in Chlamydomonas reinhardtii, one of many algae that enhance the catalytic speed of their Rubisco by packing it into a chloroplast compartment called the pyrenoid [7]. There, the algae create high local CO2 concentrations to better feed Rubisco and thus enhance CO2 fixation. On the other hand, rice and wheat, the two most consumed food crops in the world, do not have any form of CCM. So, there is growing interest in engineering a pyrenoid-based CCM into crops to improve yields while using less water and fertilizer [8].
The first step of this engineering avenue is to get an idea of the smallest number of pieces we can combine to yield a fully functional pyrenoid-based CCM. While previous works have identified essential molecular components and suggested a conceptual model for the pyrenoid-based CCM, these components remain poorly characterized, and it is unknown whether they are sufficient. To address this gap, we developed a computational model for a pyrenoid-based CCM based on the mechanism scientists believe is used by C. reinhardtii. The virtue of our modeling approach is that it allows us to systematically vary the unknown kinetic properties of each CCM component. By doing so, we were able to define a minimal set of components needed to achieve CCM function, and identify situations where together they can effectively concentrate CO2.
Below, we’ll highlight a few engineering lessons that we learned:
- In air, an engineered pyrenoid-based CCM does not require active inorganic carbon pumps, which greatly simplifies the engineering process.
- An engineered pyrenoid-based CCM can be energetically efficient, with a theoretical energy cost of only ~1.5 ATP for each CO2 This means that implementing a CCM into crop plants will likely bring more benefits than costs.
- A pyrenoid-based CCM requires a diffusion barrier to reduce pyrenoid CO2 leakage and proper enzyme localization to reduce futile cycling. Achieving both features will be key to the success of the engineered system.
Combining these engineering insights, we also proposed a stepwise engineering path to install a minimal pyrenoid-based CCM into crop plants. If you are interested in more details, please check out our paper here: https://www.nature.com/articles/s41477-022-01153-7
This work is part of an international collaboration that aims to engineer more productive crop plants by teaching them how to concentrate CO2. We hope that our model provides an initial roadmap for these ongoing engineering efforts, and serves as a useful quantitative tool to guide future research.
Reference:
- Field, C. B., Behrenfeld, M. J., Randerson, J. T., & Falkowski, P. (1998). Primary production of the biosphere: integrating terrestrial and oceanic components. Science, 281(5374), 237-240.
- Long, S. P., Marshall-Colon, A., & Zhu, X. G. (2015). Meeting the global food demand of the future by engineering crop photosynthesis and yield potential. Cell, 161(1), 56-66.
- Ray, D. K., Mueller, N. D., West, P. C., & Foley, J. A. (2013). Yield trends are insufficient to double global crop production by 2050. PloS One, 8(6), e66428.
- Leegood, R. C. (2002). C4 photosynthesis: principles of CO2 concentration and prospects for its introduction into C3 plants. J. Exp. Bot., 53(369), 581-590.
- Giordano, M., Beardall, J., & Raven, J. A. (2005). CO2 concentrating mechanisms in algae: mechanisms, environmental modulation, and evolution. Annu. Rev. Plant Biol., 56, 99-131.
- Badger, M. R., & Price, G. D. (2003). CO2 concentrating mechanisms in cyanobacteria: molecular components, their diversity and evolution. J. Exp. Bot., 54(383), 609-622.
- Meyer, M. T., Whittaker, C., & Griffiths, H. (2017). The algal pyrenoid: key unanswered questions. J. Exp. Bot., 68(14), 3739-3749.
- Hennacy, J. H., & Jonikas, M. C. (2020). Prospects for engineering biophysical CO2 concentrating mechanisms into land plants to enhance yields. Annu. Rev. Plant Biol., 71, 461-485.
Follow the Topic
-
Nature Plants
An online-only, monthly journal publishing the best research on plants — from their evolution, development, metabolism and environmental interactions to their societal significance.





Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in