To swim or not to swim during starvation
Published in Ecology & Evolution, Microbiology, and Physics
-
Motility is expensive!
"The rich are different from us”.
“Yes, they have more money”.1
I have always been fascinated by how quantitative differences can bring about qualitative distinctions. In biology, there are many examples of that. Therefore, I am a great admirer of the book “Cell Biology by the Numbers” and the associated bionumber database. During the beginning of my postdoc I was often playing with the numbers for the metabolic cost of bacterial motility, to see how this would guide their decision to swim or not. The problem was that many papers reported the cost in different units. Therefore, I started to express it in terms of ATP molecules per second, that one of my teachers described as “the 20 dollar bill in biology” (the free energy released from one ATP molecule is about 20 times the baseline energy level set by thermal noise, "kT" a fundamental energy scale equivalent to 4·10-21 J). Even though not all processes are directly driven by ATP, I find it a useful currency to quantify energy requirements.
How much does motility cost? It depends, of course, on how fast bacteria swim: many bacteria swim at about 20 μm/s, which corresponds to about 10,000 ATP/s. Many marine bacteria swim faster: at 60 μm/s, requiring 100,000 ATP/s. Is that a lot? Well, that depends. A bacterial cell requires 20 billion (20,000,000,000) ATP to double its biomass and divide (BIOID 114702). If a cell divides in 40 minutes, that’s nearly 10 million ATP/s. Compared to that, motility comprises a marginal 0.1% of cellular metabolism (or 1% if they’re swimming fast). Pondering over these numbers, I realised that during starvation, this would be completely different. Most estimates of cellular maintenance energy, the power dissipated by cells without growth, is about 10,000 ATP/s (see footnote2). So, when a bacterium swims, even at moderate speeds, it becomes one of the most expensive things a cell can do! 3
But the potential advantages during starvation are also large. When there is not enough food around you, it makes sense to start looking for more. If a bacterium does not actively move, it will diffuse, but this is really slow (it has a diffusion coefficient of about 10-8 cm2/s). A motile bacterium performs a random walk, which can be also described by diffusion, but with a 100- to a 1,000-fold higher diffusion coefficient: bringing the average search time from a year to a day. So in strongly heterogeneous environments, like the ocean, with nutrient-rich hotspots surrounded by nutrient poor waters, it is both very costly but, if a search succeeds, highly rewarding to be motile. A risk-reward trade-off was born! We started wondering how this would play out in the wild.
2. Strongly divergent behavior among relatives
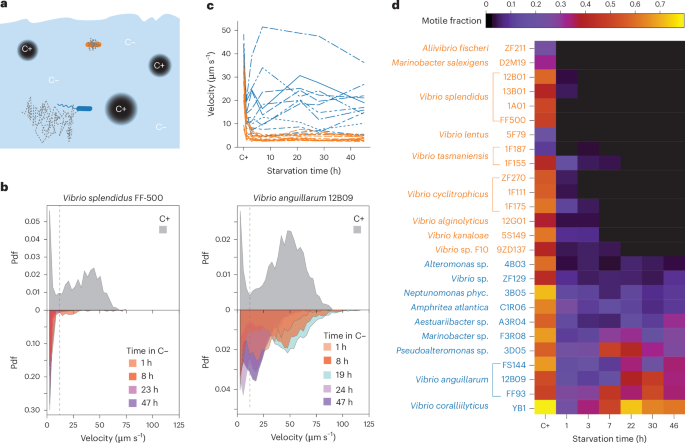
In the lab we had a collection of many marine bacteria, of which we knew some were motile and which did not like to swim (even though they had motility genes). In the literature, there seemed to be a broad consensus that starvation hinders motility, although there were reports of bacteria continuing to swim during starvation. To what extent were these differences due to differences in experiments, such as the starvation conditions or the method of quantifying motility? There was only one way to find out. This started as an undergraduate project (by Lisa Moor, who did a great job) with many restrictions in the middle of the Covid pandemic that limited lab use. We quickly found out that, when starved, closely related species behaved very differently. When starved, some species would swim, others not at all. Here is a video of a strain with an example - two bacterial species each starved for about a day, with one without any swimming cell (left) and another with many swimming cells (right):
During my PhD, all my work was performed on a single strain of a single species, E. coli RP437. I always wondered how one can say anything about nature after working on a single bacterial strain (not to say that model organisms don't have their use!). So, I really wondered if we could explore this behavior across our strain collection. We settled on 26 strains of 18 different species. I expected to find a continuum, but instead we saw that the divergent behaviors were quite general: they’d either quit motility rapidly, immediately once the washing protocol was finished, or they would swim happily for several days.
3. What's in a name?
So, there seemed to be two fundamentally different solutions to the risk-reward trade-off. It is continue searching, or stop and conserve resources. We realised it was much easier to talk about these two different behaviors if there was a name. This was a fun but slow process. I asked ChatGTP for suggestions, but these were hard to pronounce. For a while, I considered calling them “retaining” and “renouncing” strains. “Persisters” would have been perfect, but that word was already claimed for a different behavior in the context of sensitivity to antibiotics. So, reaching the limits of the English vocabulary, we had to resort to another language. In physics and mathematics there are some German words that remain in use ('Eigenvector', 'Ansatz'), but of course in biology one turns to Greek. So, I looked up an online dictionary for ancient Greek and found λιμός ('limos') for starvation, πλοῦς ('Plous') for swimming and ναφόρος ('naphoros') for moving. After pondering about the possibilities, a moment of epiphany: 'limostatic' struck me, basically combining limos (starvation) and static (well known in English, but Greek origin, too). Then the other strategy seemed straightforward: 'limokinetic'. [I have noticed, some people writing the classification for marine bacteria as 'limnokinetic', as 'limno' stands for lakes. I don’t think salinity defines our results, so not at all wrong!]
4. Bacteria as marathon-runners
I initially thought the cells that were swimming during starvation, the limokinetic strains, should die faster, but after a lot of plating and colony counting, and cell counting with flow cytometry, it was clear that the bacteria were not really dying en masse, trashing my initial hypothesis.Then, during a different experiment I noticed that the optical density of a population of swimming was going down. Of course optical density is used as a proxy for biomass. Even though I was not completely sure this was proportional, the qualitative difference to me was striking: the OD of cultures that were swimming decreased every day with about 9% of the initial biomass, while the OD of cultures that had ceased motiliy basically stayed constant.
That made a lot of sense in hindsight: for heterotrophs, the only accessible energy in this culture is the chemical energy available in carbon, and the only carbon is basically in stored in the population, in the form of biomass. (Some heterotrophs are known to be able to harvest light using rhodopsins, a different method compared to photosynthesis, but this does not play a role here, or at least not in the community we tested). With the number of cells not really decreasing, this suggested that the single-cell biomass must go down: swimming cells are sacrificing a part of their biomass to keep moving. Like marathon runners.
For some time I was proud of this project because it was my most low-tech one. In my PhD I believed the advance of science could only be achieved through technological innovation. Innovation and new instruments definitely have their place, but researchers should think of themselves as more than circus animals that perform new marvellous tricks. In any case, my pride was about to be shattered. Dieter Baumgartner, a PhD student in the lab, had been working on measuring the mass of single bacteria using holographic microscopy, an advanced microscopy technique. He came to my office, we started chatting, and he proposed to measure the mass directly. Somewhat reluctantly I agreed, because I knew that the project would not be low-tech anymore, but I now consider the result to be the strongest evidence: the swimming strain lost on average 11 fg/day worth of carbon, while the biomass of the strain that gave up motility stayed nearly constant.
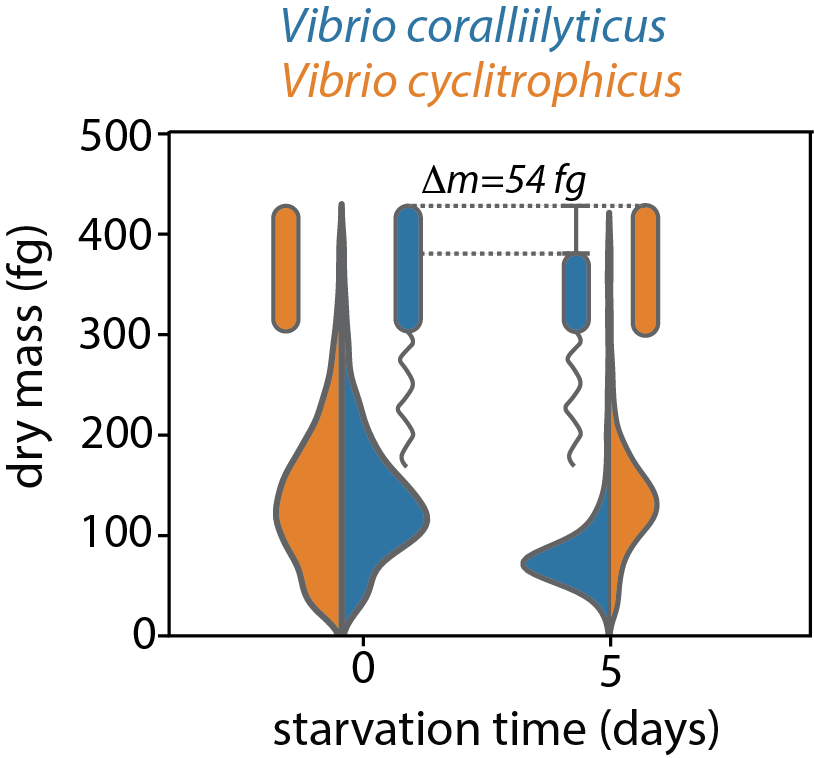
Figure: single-cell biomass distribution of a strain that stops swimming when starved (Vibrio cyclitrophicus, orange) or continues swimming when starved (Vibrio coralliilyticus, blue).
5. Motile bacteria contribute to carbon flux in the ocean
It turns out that this motile behavior is quite important in the ocean, as bacteria play a large role in controling carbon fluxes in seas and oceans. Primary producers such as phytoplankton continuously fix CO2 into biomass, feeding the marine food web. This biomass eventually starts to sink, creating a vertical carbon flux of debris called marine particles. Once it reaches the ocean interior, it can remain stored for hundreds to thousands of years, effectively sequestering carbon from the atmosphere and storing it into the ocean:
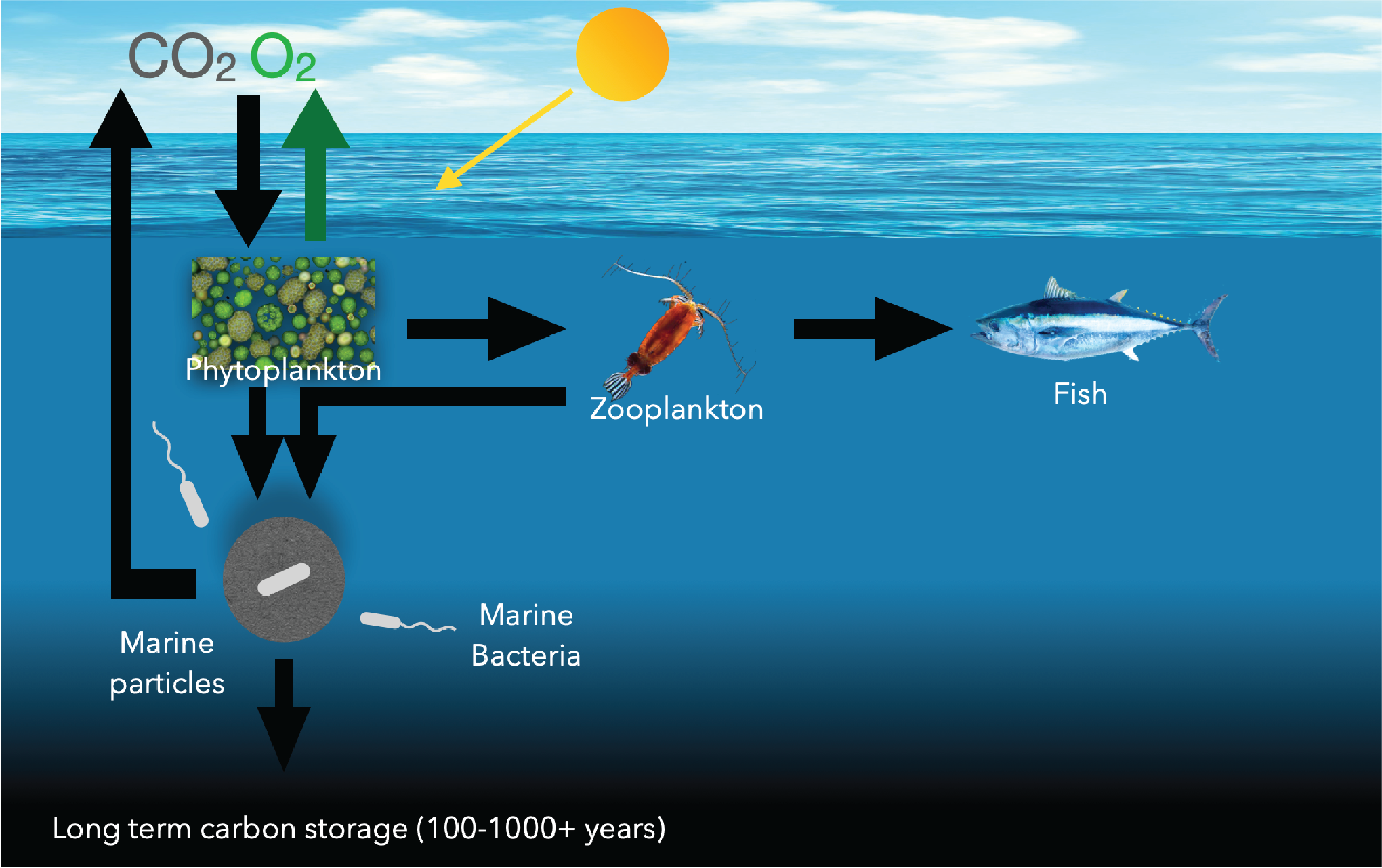
However, only a tiny fraction of the carbon that starts sinking reaches the ocean floor. For marine bacteria, these particles form nutrient-rich hotspots that they can degrade and thereby release the stored carbon back to the atmosphere, effectively acting as gatekeepers of carbon storage. Since these particles are dilute, it is a challenge for bacteria to find them and therefore understanding this process clearly does not only depend on the biological adaptations to life on such particles, but also the challenge in finding such particles. Motility is a great way to enhance the finding of such particles, but the concentrations of nutrients outside of these particles are so low that motile bacteria (there are many bacteria outside of hotspots, but these bacteria are adapted to life outside of the particles). So, it is important to understand under which conditions bacteria are motile and if they can continue searching for particles even if the environment does not provide sufficient background energy to continue swimming.
6. Using genomic predictions to connect lab to field.
One way to generalise predictions from a lab to the environment, in our case the ocean, is to make a genetic classifier. I had no clue about bioinformatics, but luckily there were people that were willing to help with this. Zach Landry used machine learning classifers to find a set of genes that could separate the two groups with nearly 90% accuracy, and we were able to predict the outcome for strains that we had not used for training with about 85% accuracy. I certainly do not take this result for granted. There were many reasons why this may not have worked!
A genomic classifier allows us to make predictions in the field, based on a limited set of observations in the lab. Some of the strains used in our study were isolated from the same area, indicating that these strategies can co-exist in the same environments. But certainly environmental factors could influence which strategy is more favourable when and where. The classifier predicts that overall, in the ocean the limokinetic strategy dominates. However, there seem to be areas where the limostatic strategy dominates. We can’t wait to test such predictions in the field!
7. Conclusion
With the daunting diversity of microbial life, it is gratifying to stumble upon some simplicity behind it. That discovery was a great journey and I thank all my co-authors for their invaluable contributions (there were many other important contributions that I could not highlight here because of length reasons). I am also grateful for all the insights I obtained from people asking a question at a poster or seminar presentation!
Footnotes
1) I borrow this example from P.W. Anderson's seminal paper "More is different" (Science, 1972).
2) A good source is Biselli et al. MSB 2020. It's given as 0.61 mmol ATP/h/g, I assume a dry weight of 150 fg per cell. It can be ~2 fold higher or lower depending on the growth rate before starvation.
3) For more numbers and references, see our review "The ecological roles of chemotaxis" (Nat. Rev. Microb., 2022)
Image credit for banner picture: Sophie Zweifel and Pallavi Keshri , ETH Zurich
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in