
Understanding Multicellularity in Streptomyces
Historically much of our understanding of the mechanisms that underpin cell division in bacteria has been driven by studies using classical model organisms such as Escherichia coli and Bacillus subtilis. In recent years however, research from labs using “non-classical” organisms has revealed the tremendous diversity in the regulation and cell biology of division within the microbiological landscape.
The abundant soil bacteria Streptomyces are a case-in-point. Although they are bacteria, filamentous growth of thread-like hyphae occurs to create a branching, compartmentalised mycelium. Indeed, their complex growth and morphology led early microbiologists to believe they were in fact eukaryotic fungi. Instead, as we now know, bacteria can either grow as unicellular or multicellular organisms. Multicellularity in bacteria represents an important step in evolution across the domains of life (Lyons and Koulter 2015). Although there is an energetic cost for Streptomyces, there are numerous advantages to a multicellular lifestyle. Likely benefits include increased resistance to environmental stresses and protection in the competitive soil environment, improved gathering of nutrients and the opportunity for “division of labor” in which tasks are attributed across different cell-types (Zhang et al. 2016; West and Cooper 2016).
What makes Streptomyces a fascinating model for bacterial cell division is that when nutrients are scarce, the multicellular mycelium undergoes a coordinated program of differentiation to form unicellular spores. After being dispersed, these spores germinate to re-establish a multicellular mycelium in a new environment.
All this means that unlike most unicellular bacteria which divide by binary fission to yield two virtually identical daughter cells, two distinct modes of cell division exist in Streptomyces:
- Cross-Wall Formation During the vegetative growth phase, division septa also called “cross-walls” partition the hyphal mycelium into a series of connected multi-genomic compartments.
- Sporulation-specific cell division During reproductive growth, ladder-like arrays of septa divide the multi-genomic hyphae into uni-genomic spores.
At the heart of both modes is the bacterial tubulin-homolog FtsZ that forms the contractile Z-ring that acts as a scaffold for the assembly of the other division components that collectively form a large protein division machinery, the so-called “divisome”. As part of the Schlimpert Lab (John Innes Centre, Norwich, UK), I am interested in understanding the mechanisms that drive both modes of cell division in Streptomyces.
At the outset, our research was centred around two main questions:
- How is multicellularity established in Streptomyces and why is it important? Although cross-walls have long-since been observed, how they are formed has remained unknown. Up until this point, no protein (apart from FtsZ-itself) had been identified to play a role in cross-wall formation.
- How does Streptomyces navigate the additional complexity involved during sporulation-specific cell division? Sporulation-specific cell division in Streptomyces involves significant complexity that does not feature in unicellular bacteria. Rather than placing a single Z-ring at mid-cell Streptomyces must correctly place dozens of Z-rings in multi-nucleate hyphae of no fixed length so that each resulting spore carries a single copy of the chromosome. Importantly, the canonical control mechanisms that mediate proper segregation and septation in unicellular bacteria are entirely absent in Streptomyces, suggesting that alternative systems exist.
The identification of SepX
To address these questions, I went in search of novel components of the Streptomyces divisome important for cross-wall formation and sporulation. Previously, I had spent time defining the regulatory networks that underpin the Streptomyces developmental cascade. Using Chromatin Immunoprecipitation-sequencing (ChIP-seq) I identified all the genes under the control of two key transcription factors (WhiA and WhiB) that are essential for sporulation in Streptomyces (Bush et al., 2013; Bush et al., 2016). WhiA and WhiB co-activate the transcription of several genes encoding core members of the bacterial cell division machinery including ftsZ but we reasoned that other genes important for cell division might also be under WhiAB-control. I constructed and screened a series of strains expressing fluorescently tagged WhiAB-target proteins, searching for those that co-localise with FtsZ at future sites of cell division. This is how we came to identify sepX, encoding a novel component of the divisome in Streptomyces required for compartmentalisation and sporulation.
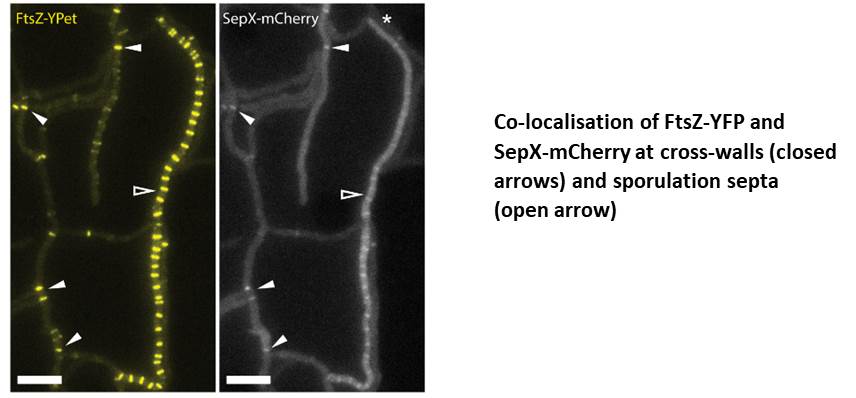
SepX is essential for cross-wall formation, allowing us to study the importance of multicellularity
It was an exciting moment when I first noticed that deletion of sepX leads to a complete loss of cross-walls and multicellularity during vegetative growth, leading to giant filamentous cells that shared a continuous cytoplasm. This was a landmark finding since SepX turned out to be the first component of the division machinery (apart from FtsZ itself) that is specifically required for cross-wall formation.
Importantly, the failure of the sepX mutant to synthesise cross-walls allowed us to finally assess the importance of hyphal compartmentalisation for the growth and fitness of Streptomyces. In contrast to the whiA mutant (that forms cross-walls but lacks sporulation septa), the sepX mutant (that forms sporulation septa but lacks cross-walls) has a significant fitness defect. sepX mutant colonies are much smaller and grow much slower than either the whiA mutant or the wildtype. A lack of hyphal compartmentalization in the ftsZ (Santos-Beneit et al., 2017) and sepX mutants increases the propensity of lysis in growing hyphae, in line with the idea that cross-walls protect the mycelial mass from large-scale loss of viability. In the absence of cross-walls we further showed that FtsZ-ladder formation during sporulation is not restricted to individual hyphal tip compartments but occurs across the entire mycelial network, suggesting that the organisation of the mycelium into compartments is important for the ordered progression through the lifecycle. It is possible that the accumulation of molecular signals or regulators is particularly vital. In the absence of cross-walls, an inbalance in the concentrations of these key factors may therefore impair the transition to the next step in the developmental program.
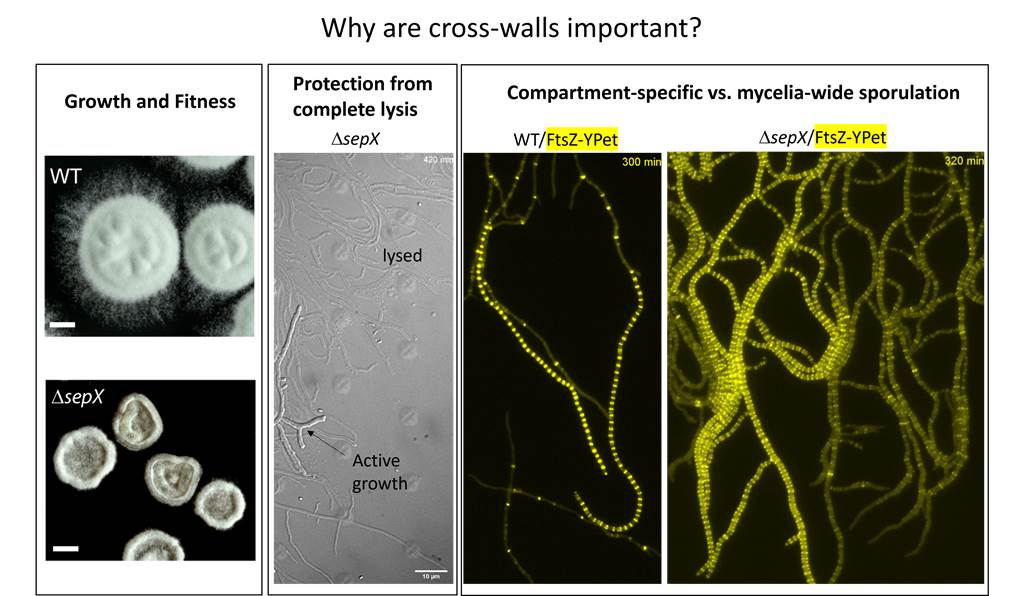
A key role for SepX during sporulation-specific cell division
Although the sepX mutant does not form cross-walls, it does sporulate. However, spore septa are irregularly spaced and often placed in an altered plane leading to irregularly sized spores. By employing time-lapse microscopy to track the movement of YPet-tagged FtsZ in real-time we were able to show that in the sepX mutant the assembly dynamics and the architecture of Z-rings was altered. Compared to the wildtype, Z-rings showed reduced fluorescent intensity and were wider. This suggests that SepX is important for the spatial stabilisation of Z-rings, and ultimately septum formation.
Since SepX is a membrane protein with no obvious enzymatic function, we reasoned that it is likely to perform a scaffolding function, bringing other key components of the division machinery into play. We found that SepX co-purifies and interacts with the sporulation-specific cell division component DynB (Dynamin-like protein B). We previously showed that DynB functions to stabilise Z-rings during sporulation via further interactions with other division components (Schlimpert et al., 2017). While both sepX and dynB single mutants do sporulate, a sepXdynB double mutant forms very few viable spores. This underlines the collective importance of SepX and DynB in the scaffolding of the divisome and stabilisation of Z-rings during sporulation.
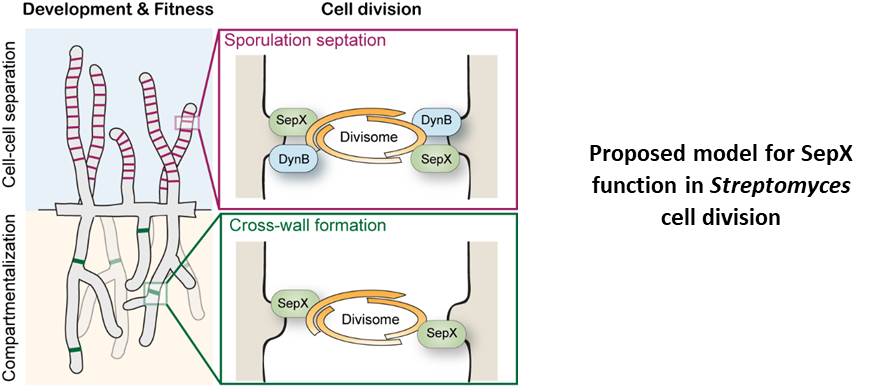
Multicellularity and cell-cell communication
Streptomyces differentiation is dependent upon the proper expression of key transcriptional regulators and the appropriate concentration of signalling molecules such as cyclic-di-GMP (Bush et al., 2015). The near-synchronous nature of transition from vegetative to reproductive growth suggests that the series of interconnected compartments created by cross-walls are unlikely to function autonomously and that it is important for signals and nutrients to be shared across the mycelium both during growth and upon differentiation. Cell-cell communication channels have been widely reported in other multicellular organisms, including plasmodesmata in plants, septal pores in fungi and septal junctions in filamentous cyanobacteria, raising the possibility that such structures may exist within vegetative cross-walls. Indeed, a recent structural study suggests the presence of a pore-like structure in Streptomyces cross-walls (Sexton and Tocheva 2020). Our discovery of sepX, may therefore help the identification and study of such channels in filamentous actinobacteria.

References
Bush M., Bibb M., Chandra G., Findlay K. and Buttner M. Genes required for aerial frowth, cell division, and chromosome segregation are targets of WhiA before sporulation in Streptomyces venezuelae. mBio 4, e00684-13 (2013).
Bush M., Tschowri N., Schlimpert S., Flärdh K. and Buttner M. c-di-GMP signalling and the regulation of developmental transitions in streptomycetes. Nat Rev Microbiol. 13(12):749-60 (2015).
Bush M., Chandra G., Bibb M., Findlay K. and Buttner, M. Genome-wide chromatin immunoprecipitation sequencing analysis shows that WhiB is a transcription factor that cocontrols its regulon with WhiA to initiate developmental cell division in Streptomyces. mBio 7, e00523-16, (2016).
Lyons N. and Kolter R. On The Evolution of Bacterial Multicellularity. Curr Opin Microbiol. 24: 21–28.(2015)
Santos-Beneit F., Roberts D., Cantlay S., McCormick J. and Errington J. A mechanism for FtsZ-independent proliferation in Streptomyces. Nature Comms 8: 1378 (2017)
Schlimpert S., Wasserstrom S., Chandra G., Bibb M., Findlay K., Flärdh K. and Buttner M. Two dynamin-like proteins stabilize FtsZ rings during Streptomyces sporulation. Proc Natl Acad Sci USA 114, E6176–E6183 (2017)
Sexton D and Tocheva E. Ultrastructure of Exospore Formation in Streptomyces Revealed by Cryo-Electron Tomography. Front Microbiol, 11:581135. (2020)
West S. and Cooper G. Division of labour in microorganisms: an evolutionary perspective. Nat Rev Microbiol. volume 14: 716–723 (2016)
Zhang Z., Claessen D. and Rozen D. Understanding Microbial Divisions of Labor. Front Microbiol 7:2070. (2016)
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in