Unraveling the Marvels of Endothelial SMAD1/5 in Juvenile Long Bones
Published in Cell & Molecular Biology and Anatomy & Physiology
I would like to use this blog post to share some personal insights, how this project was inspired and how it evolved to become such an exciting international collaboration project across 2 continents and 3 countries. I hope this serves as a wonderful example to show that marvelous science and unexpected findings can arise when you follow your curiosity for biological systems, your instincts as scientists and your motivation to engage with experts from different fields.
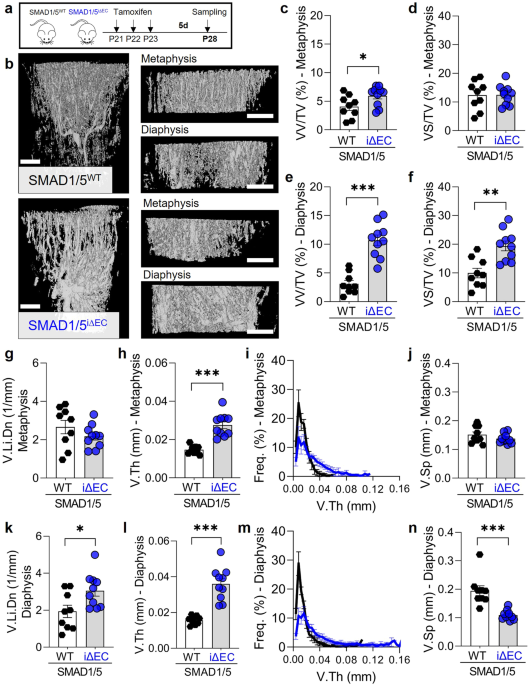
The crosstalk between the vasculature and bone has fascinated many generations of researchers, but several questions still remain open. Osteoblasts, as bone-forming cells, require close proximity to blood vessels leading to the assumption that in bone, angiogenesis and osteogenesis are coupled. During bone development, the primary spot for bone formation and angiogenic-osteogenic coupling is in the growth plate – putting a spotlight on metaphyseal vessels ending at the chondro-osseus junction to support and nurture the powerhouse of bone growth (Figure 1). In 2014, a team of scientists – Anjali P. Kusumbe, Saravana K. Ramasamy, Ralf H. Adams precisely characterized the metaphyseal vessels in the murine skeletal system during post-natal development directing the coupling of angiogenesis to osteogenesis [1]. The Notch signaling pathway was identified as major regulator of this coupling process [2].
This amazing team of scientists and their publications (and many which followed afterwards) inspired a whole range of new studies in bone research and also draw my interest as a PhD student at the Charité-Universitätsmedizin Berlin, Germany and member of the Berlin-Brandenburg School for Regenerative Therapies (BSRT). My research work focused on developing and testing a new regenerative therapy to improve delayed bone fracture healing [3]. Successful bone regeneration upon fracture strongly depends on the speed and completeness of reestablishing a functional vascular network to ensure nutrient supply and proper mineralization at the callus site. Hence, revascularization in the fracture callus depends on a functional coupling of angiogenesis to osteogenesis. Interestingly, newly formed vessels at the forefront of the fracture callus show great similarities with the metaphyseal vessel subtype described during post-natal development, supporting the concept that natural fracture healing is most efficient when it recapitulates development. Thus, dissecting the underlying mechanism of angiogenic-osteogenic coupling advances our understanding of developmental processes and fracture repair. The therapeutic of interest that I was focusing on during my PhD particularly accelerated revascularization in the callus area. Therefore, I became excited to learn more about the interaction between bone formation and the vasculature. However, at this stage of my career, I had no particular knowledge of vascular development or sprouting angiogenesis, or any potential underlying pathways such as Notch or bone morphogenic proteins (BMP).
Thanks to my graduate program, I had a great peer network across disciplines. Andreas Benn, a fellow PhD student (and now partner for life), became a BMP expert during his PhD and joined An Zwijsen's group at KU-Leuven, Belgium during his Postdoc in 2016. There he focused on the role of endothelial SMAD1/5 activity in neurovascular development in the murine retina [4]. SMAD1/5 are downstream effectors of BMP which determine the final functional outcome of the upstream signal. It was during one of our typical discussions that Andy tried to explain the whole Notch vs. BMP/SMAD signaling pathways in vascular angiogenesis to me when we recognized that it was unknown whether endothelial SMAD1/5 activity plays a role in BMP-regulated coupling of osteo- and angiogenesis. This is of special interest since BMP is the major regulator of bone development, formation, and homeostasis – meaning developing bone contains a lot BMPs. However, it is unknown how this BMP-rich environment affects the surrounding vasculature, for example, through SMAD1/5. As Andy was working with endothelial specific SMAD1/5 knockout mice and only required the retinas for his research, he started to send me some bone samples just to have a look. Screening the first samples with histology made me wonder – especially the diaphyseal vasculature was tremendously disrupted (Figure 2) and the growth plate looked different than in wild type mice.
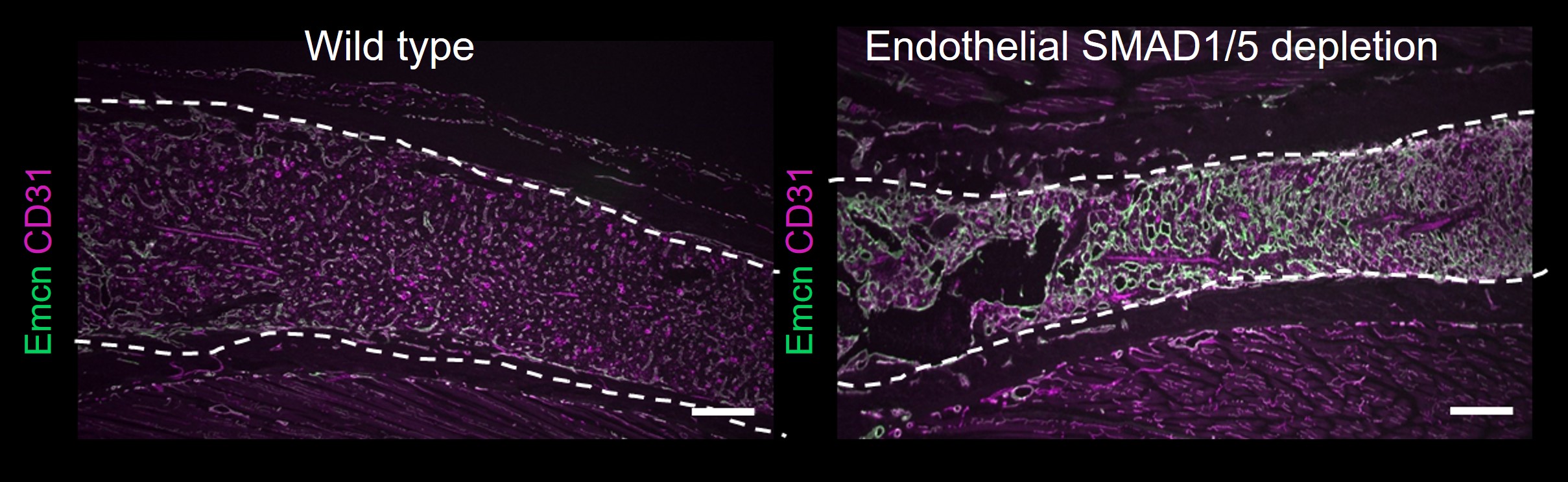
Figure 2: Immunofluorescence images show diaphyseal vasculature in bone samples from juvenile mice – comparing control samples (wild-type) and samples from mice with endothelial SMAD1/5 depletion. Staining indicates the vessel marker CD31 and endomucin (Emcn).
These initial findings motivated me to visit KU Leuven in 2019 for a few weeks to work closely with An Zwijsen and to perform high-resolution contrast-enhanced computer tomography (NanoCT) together with Greet Kerckhofs. The aim was to visualize the vasculature in 3D and investigate the properties of the bone (Figure 3). Using this powerful technique, we found that endothelial-specific SMAD1/5 depletion resulted in larger, dilated metaphyseal and diaphyseal vessels (Figure 3) and alterations in the trabecular, growth plate adjacent, and cortical bone formation area.
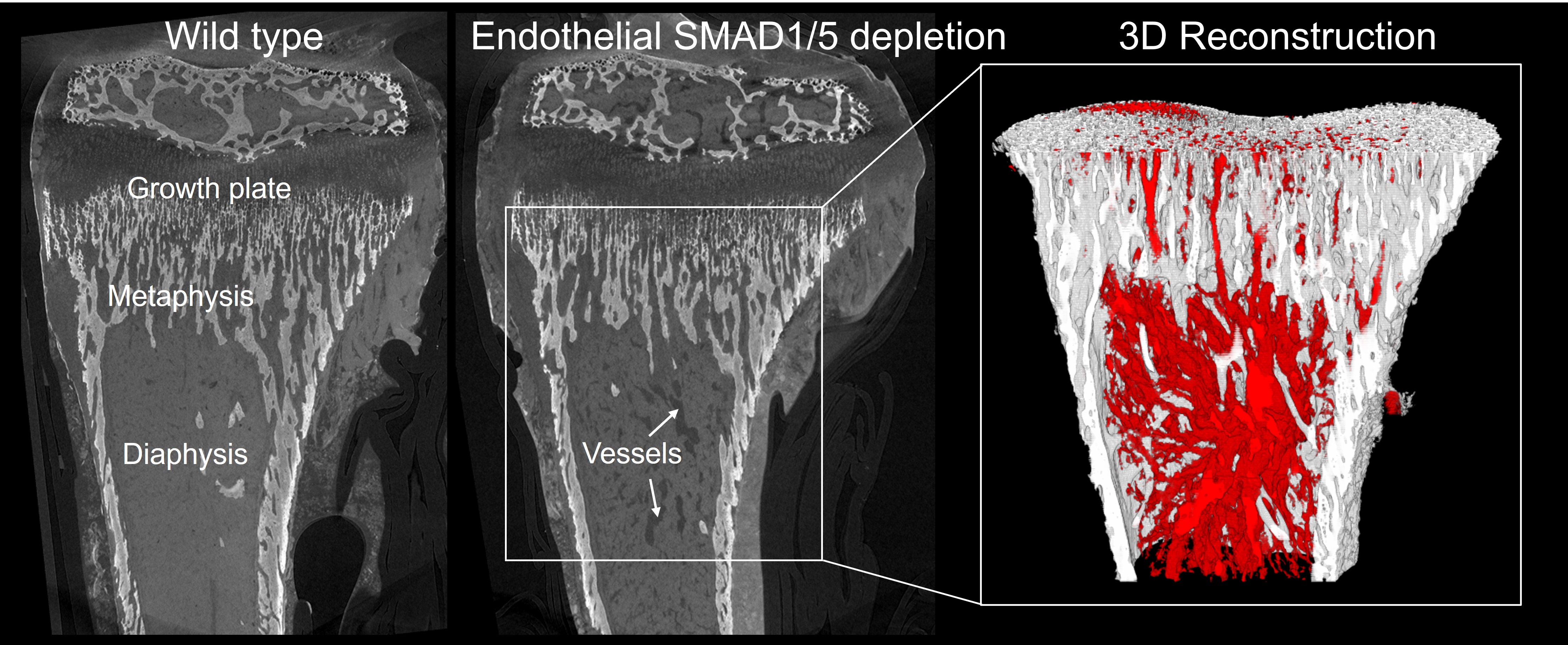
Figure 3: We performed high-resolution contrast enhanced computer tomography on control samples (wild-type) and samples from mice with endothelial SMAD1/5 depletion. Images on the left show exemplary sections from the scans and anatomical descriptions – vessels are stained in dark grey. Image on the right side is an example for a 3D reconstruction marking bone in white and blood vessels in red in the samples with SMAD1/5 depletion highlighting the enlargement of the vessels in the bone marrow channel.
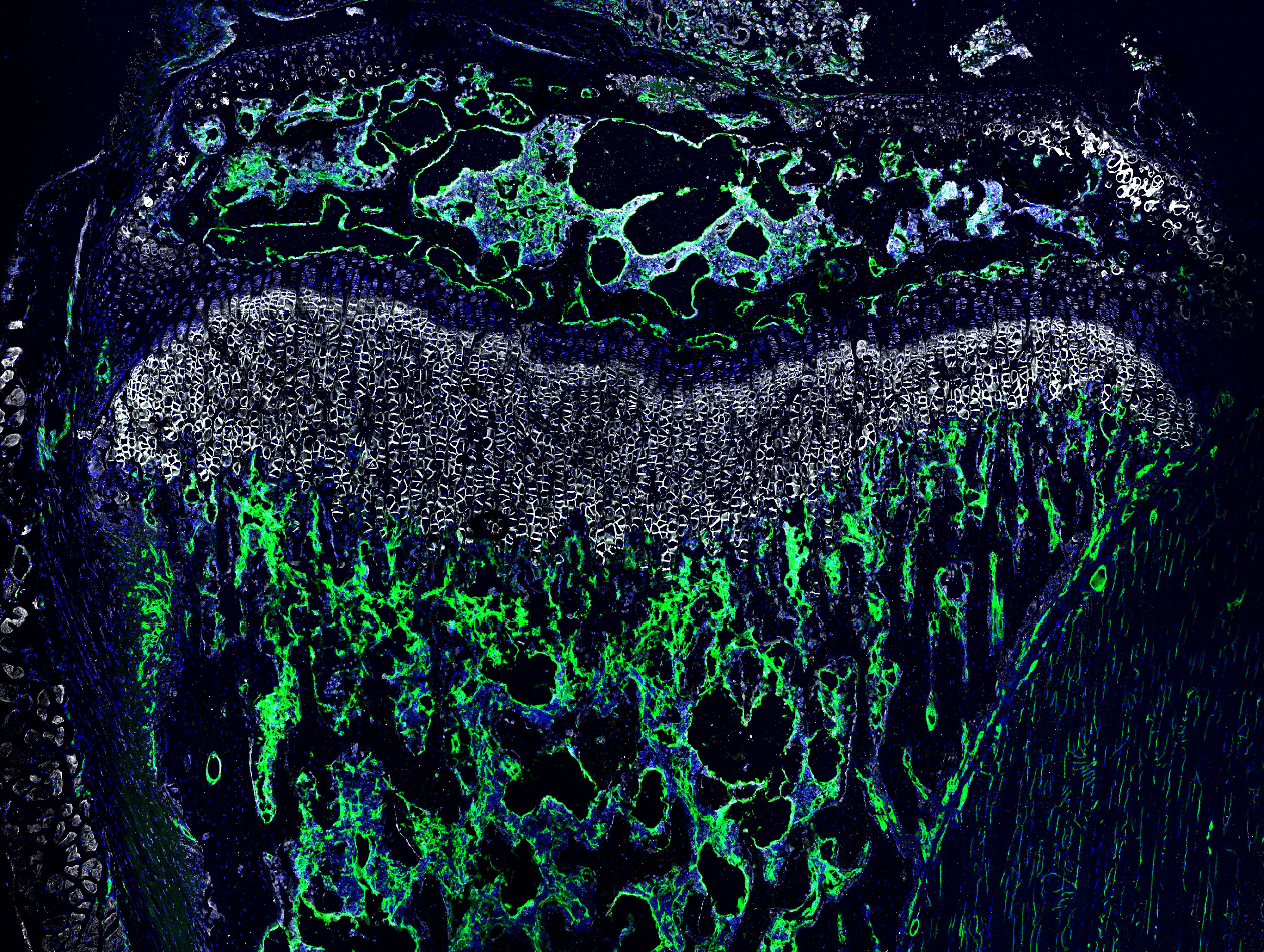
In 2019, I met Joel D. Boerckel at the ORS Annual Meeting in Austin, Texas and his scientific curiosity and wonderful personality, convinced me to join his lab as a Postdoc. Joel was immediately excited about this project and more than willing to help me to fill the gaps in the understanding and storyline. In 2021, when I joined his lab at the University of Pennsylvania in Philadelphia, I brought a box full of samples – turning the project into an international collaboration across 2 continents and 3 countries. Together, Joel and I were able to show that endothelial SMAD1/5 activity maintains skeletal vascular morphogenesis and function in the metaphysis and particularly the diaphysis, inducing not only larger loop formation but also elevating vascular permeability (more detailed analysis of previous findings highlighted in Figure 2 and 3). In addition, growth plate resorption was impaired which resulted in an enlarged growth plate thickness (Figure 4; upper panel) accompanied by an inhibition of osteoprogenitor recruitment to the primary spongiosa upon long endothelial SMAD1/5 depletion (Figure 4; lower panel; no OSX-expressing cells with SMAD1/5 depletion).
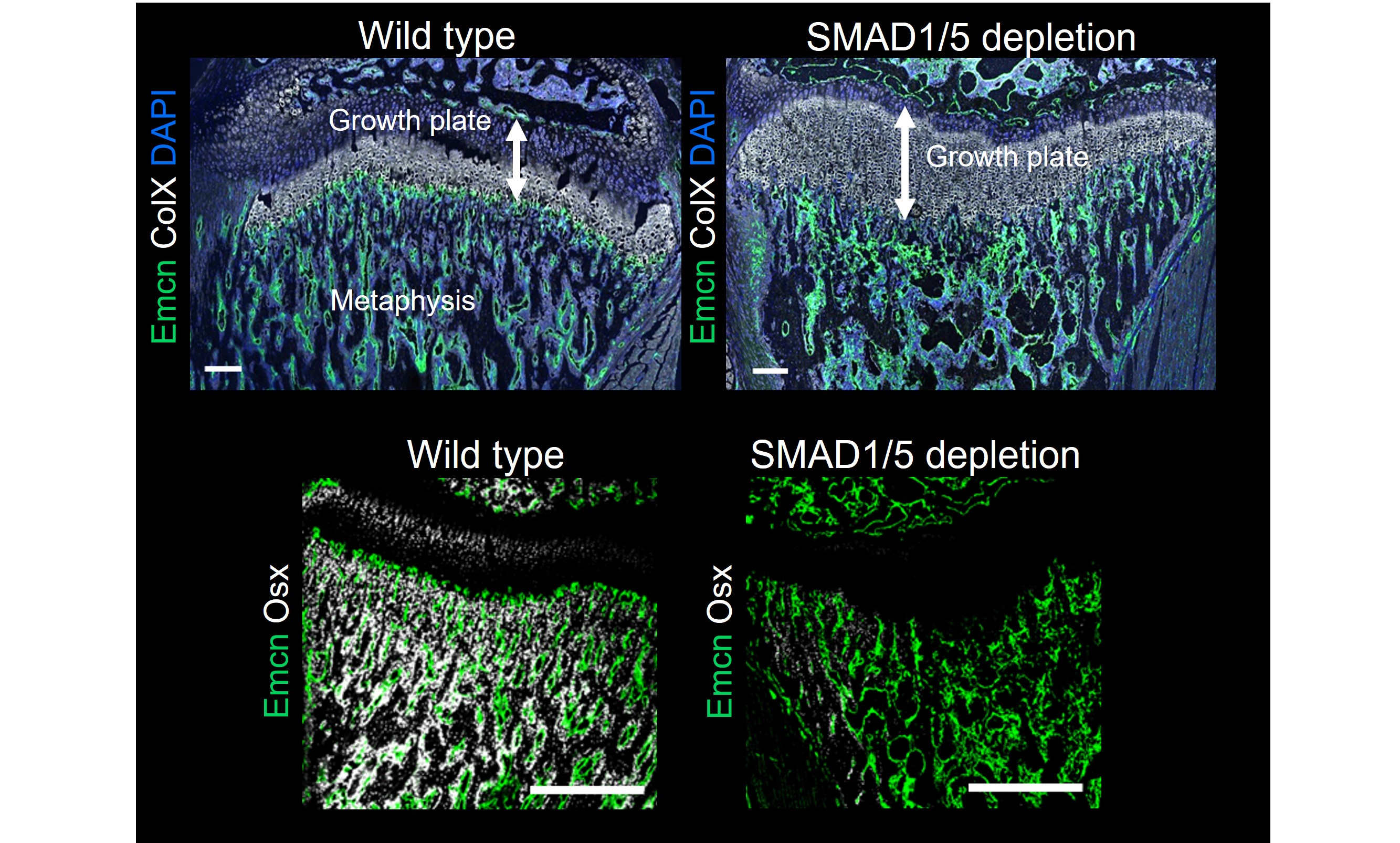
Figure 4: Immunofluorescence images show changes in the growth plate and metaphyseal vasculature (upper panel) and metaphyseal bone formation area (lower panel) in bone samples from juvenile mice – comparing control samples (wild-type) and samples from mice with endothelial SMAD1/5 depletion. Staining indicates the vessel marker endomucin (Emcn), the growth plate marker collagen 10 (ColX), the cell nuclei marker DAPI and the osteoblast marker osterix (Osx).
These findings, particularly the effects on the diaphyseal vasculature will be exciting to further understand during the next steps of my career – as the vascular phenotype shows similarities to what is happening in the bone marrow surrounding the fracture gap. During bone injury and subsequent regeneration program, the diaphyseal vasculature requires an impressive flexibility and plasticity to ensure nutrient and oxygen supply. How likely is it that BMP/SMAD1/5 signaling is involved in this process?
This journey exemplifies the beauty of scientific collaborations across borders, and I am more than grateful for all the amazing scientists and mentors I was able to work with and who selflessly supported me in finalizing this project – THANK YOU Andy, An, Greet & Joel. Our findings not only shed light on the role of endothelial SMAD1/5 in mediating angiogenic-osteogenic coupling but also pose intriguing questions for future exploration.
Graphical summary:
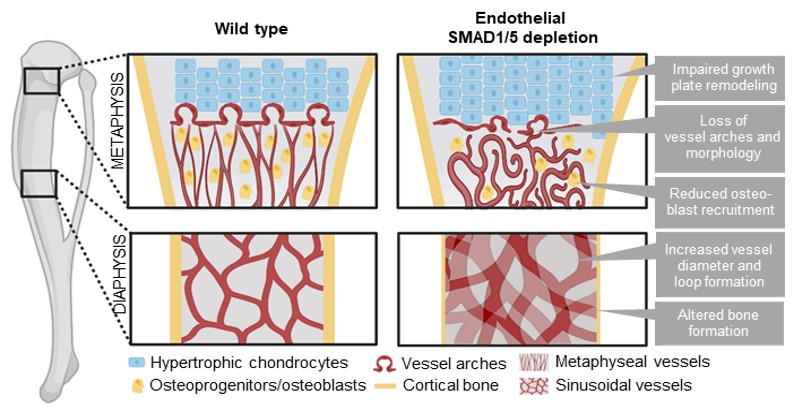
Figure 5: Graphical summary on effects of endothelial SMAD1/5 depletion on metaphyseal and diaphyseal vessel and bone formation during juvenile and early adolescent long bone growth. Illustration has been created with BioRender.com.
Link to the paper: https://rdcu.be/dA4Q1
References:
[1] Kusumbe, A. P., Ramasamy, S. K. & Adams, R. H. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nature 507, 323-328, (2014). https://www.nature.com/articles/nature13145
[2] Ramasamy, S. K., Kusumbe, A. P., Wang, L. & Adams, R. H. Endothelial Notch activity promotes angiogenesis and osteogenesis in bone. Nature 507, 376-380, (2014). https://www.nature.com/articles/nature13146
[3] Lang, A. et al. MIF does only marginally enhance the pro-regenerative capacities of DFO in a mouse-osteotomy-model of compromised bone healing conditions. Bone 154, 116247 (2022). https://www.sciencedirect.com/science/article/abs/pii/S8756328221004130?via%3Dihub
[4] Benn, A. et al. BMP-SMAD1/5 Signaling Regulates Retinal Vascular Development. Biomolecules 10, 488 (2020). https://www.mdpi.com/2218-273X/10/3/488
Follow the Topic
-
Communications Biology
An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.

Related Collections
With Collections, you can get published faster and increase your visibility.
Artificial Intelligence Methodology in Structural Biology
Publishing Model: Hybrid
Deadline: Nov 30, 2026
Healthy Aging
Publishing Model: Open Access
Deadline: Dec 31, 2026

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in