2018: The year of bacterial anti-phage defenses?
Published in Microbiology
Man, her last work, who seem'd so fair,
Such splendid purpose in his eyes,
…
Who trusted God was love indeed
And love Creation's final law—
Tho' Nature, red in tooth and claw
With ravine, shriek'd against his creed—
- Alfred Tennyson, In Memoriam A.H.H., Canto 56
The war between predator and prey, between bacteria and their phages is perhaps as old as life itself. Bacteria, as prey, have an arsenal to defend themselves against their ancient predators and phage, in their turn, are just as ingenious in developing engines to breach these defenses. Diverse bacterial anti-phage determinants have been discovered and described in great detail. These include not only the well-known restriction-modification and CRISPR-Cas systems, but also BREX1 (bacteriophage exclusion), Abi2 (abortive infection), Pgl3–5(phage growth limitation) and prokaryotic argonaute-like proteins6.
In March 2018, Doron et al.7 reported nine distinct functional anti-phage systems and one anti-plasmid system after a large-scale bioinformatics analysis of prokaryotic genomes followed by testing for function in E. coli or B. subtilis. Obviously, due to the limitations of heterologous hosts for functional analyses, the actual number of defense systems might be even larger. In October 2018, Ofir et al.8 reported the widespread existence of DISARM7 (‘defense island system associated with restriction-modification’) in prokaryotic genomes and functionally tested one such system derived from Bacillus paralicheniformis by cloning it into B. subtilis. Now, to conclude this year, a detailed report9 (in some ways also detailed and thorough reappraisal) has been published by Kronheim et al. in December 2018 describing chemical defenses against phage infection in the Streptomyces. They specifically focus on the production of anthracyclines – a class of DNA-intercalating molecules – of which doxorubicin, daunorubicin, epirubicin and idarubicin exhibit antibiotic properties and are used in cancer therapy as well.
Studies of the anti-phage properties of some of these Streptomyces secondary metabolites go back more than fifty years10,11. However, in their current paper, Kronheim et al.9 bring the power of high-throughput metabolomics to bear upon the problem and identify several such molecules produced by Streptomyces. Very importantly, they have determined that the anthracyclines do not inhibit viral attachment or DNA injection. Rather anthracyclines inhibit some process occurring after injection of viral DNA into the host cell, but before viral replication. Of renewed interest is their verification that sub-inhibitory concentrations of daunorubicin in the growth medium can protect E. coli and Pseudomonas aeruginosa from phage infection, as determined by the reduction in phage titer post-infection (Table 1 in Kronheim et al.). A notable observation in this work is that this protection appears to be specific to infection by double-stranded DNA phages, and does not to extend to protection against the single-strand DNA phage M13 for E. coli.
Now, given that bacteria form part of a community and are seldom, if ever, encountered as pure cultures, this work has highlighted yet again the value of common goods for community establishment and maintenance. Being diffusible molecules, anthracyclines are capable of entering other bacterial cells and, at sub-inhibitory concentrations, confer anti-phage protection on such lucky cells. Therefore, the spatial arrangement of microbiota as biofilms on solid substrates – whether teeth or soil – may be significantly modulated and maintained by the presence or absence of Streptomyces-like species. The production of secondary metabolites is a mechanism that keeps nearby microbial competitors (and phages) at bay (at inhibitory concentrations) but benefits more distant bystanders (at lower concentrations) by supplementing their phage defenses. A few micrometers perhaps determine the difference between life and death when other bacteria are in the vicinity of Streptomyces. Could this be termed herd immunity at a (safe) distance?
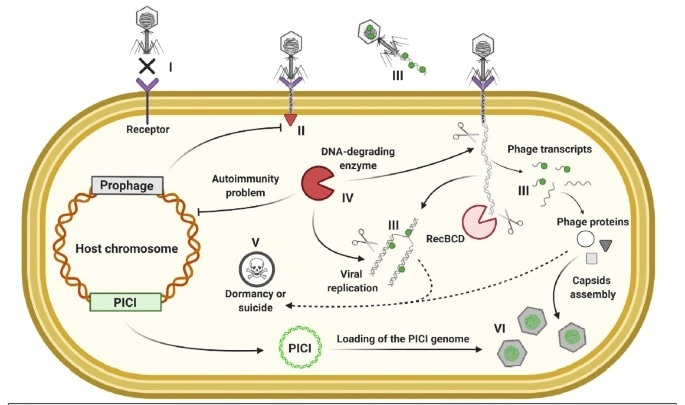
Poster image from Fig. 1 in Isaev, A.B., Musharova, O.S. & Severinov (2021)13 under the Creative Commons Attribution 4.0 International License.
References:
1. Goldfarb, T. et al. BREX is a novel phage resistance system widespread in microbial genomes. EMBO J. 34, 169–183 (2015).
2. Dy, R. L., Przybilski, R., Semeijn, K., Salmond, G. P. C. & Fineran, P. C. A widespread bacteriophage abortive infection system functions through a Type IV toxin-antitoxin mechanism. Nucleic Acids Res. 42, 4590–4605 (2014).
3. Chinenova, T. A., Mkrtumian, N. M. & Lomovskaia, N. D. Genetic characteristics of a new phage resistance trait in Streptomyces coelicolor A3(2). Genetika 18, 1945–1952 (1982).
5. Hoskisson, P. A., Sumby, P. & Smith, M. C. M. The phage growth limitation system in Streptomyces coelicolor A(3)2 is a toxin/antitoxin system, comprising enzymes with DNA methyltransferase, protein kinase and ATPase activity. Virology 477, 100–109 (2015).
6. Swarts, D. C. et al. DNA-guided DNA interference by a prokaryotic Argonaute. Nature 507, 258–261 (2014).
9. Kronheim, S. et al. A chemical defence against phage infection. Nature 564, 283–286 (2018).
10.Parisi, B. & Soller, A. Studies on the antiphage activity of daunomycin. Giorn. Microbiol. 12, 183–194 (1964).
11.Morita, J., Tanaka, A., Komano, T. & Oki, T. Inactivation of Phage ϕ X174 by Anthracycline Antibiotics, Aclacinomycin A, Doxorubicin and Daunorubicin. Agric. Biol. Chem. 43, 2629–2631 (1979).
Disclaimer: The opinions expressed herein do not represent the views of the TERI School of Advanced Studies or TERI.




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in