An Aspartimide Containing Graspetide RiPP
Published in Chemistry
Have you ever felt frustrated by weeds in your garden? No matter how much you try to keep your garden clean and beautiful, these weeds are a nuisance. Now, what if I told you that these weeds had a purpose? They were meant to be there. That is exactly what we found of the aspartimide in our newly characterized ribosomally synthesized and post-translationally modified peptide (RiPP), fuscimiditide!
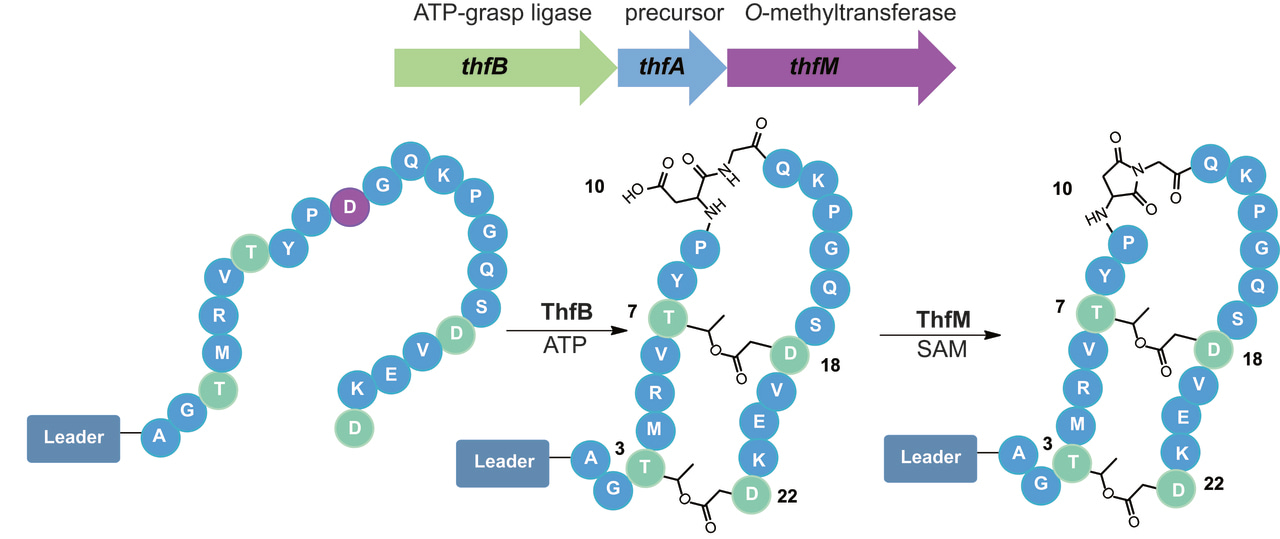
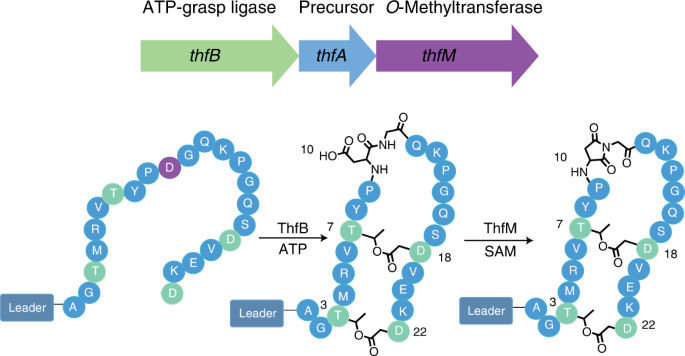
Over the course of our bioinformatic search for unique RiPPs in 2017, we came across an interesting genome. This genome had a gene cluster with all the distinguishing features of graspetides, lactone/lactam containing macrocyclic RiPPs [1], yet it had an additional gene that had yet to be explored. This gene was annotated as an O-methyltransferase which is reminiscent of protein repair enzymes called PIMTs. These PIMTs usually correct isoaspartates that have been formed due to protein aging back to their native aspartate form. This repair pathway proceeds through an aspartimide intermediate which allows for this repair to occur (Figure 1). This resemblance to PIMTs intrigued us and piqued our interest to study this gene cluster found in Thermobifida fusca.
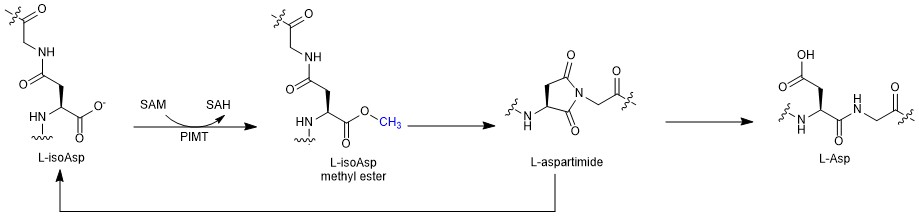
Figure 1. PIMT repair pathway. Over time, protein aging can cause aspartic acid (Asp) to isomerize to isoaspartic acid (isoAsp). This noncanonical isoAsp residue can be repaired by PIMT which uses S-adenosyl methionine (SAM) to create a methyl ester that is readily hydrolyzed to an aspartimide intermediate. Hydrolysis of the intermediate can result in restoration of Asp to its canonical state.
Through heterologous expression of the graspetide-like genes and the O-methyltransferase in E. coli, the data suggested that the enzyme was catalyzing an extra dehydration. For a while, we debated whether the dehydration was due to an amide linkage or aspartimide formation. We could not fathom that nature would purposefully install what is otherwise a nuisance side product in peptide synthesis or an unstable intermediate in a protein repair pathway. Yet, through tandem mass spectrometry, hydrazine reactions, and most importantly, NMR studies, we were able to conclusively attribute this extra dehydration to aspartimide formation (Figure 2). This O-methyltransferase also had promiscuous activity; not only did it modify Asp to an aspartimide, but it could also modify isoAsp to an aspartimide as well! While the whole story is too grand to cover in this post, some of the other highlights include: 1) fuscimiditide’s precursor has a really long leader peptide and most of it is required for ester formation by ThfB, 2) the esters must form first before aspartimidylation can occur, 3) ThfM can catalyze aspartimidylation whether the leader peptide is present or not, and 4) precursor modification can be done in vivo and in vitro.
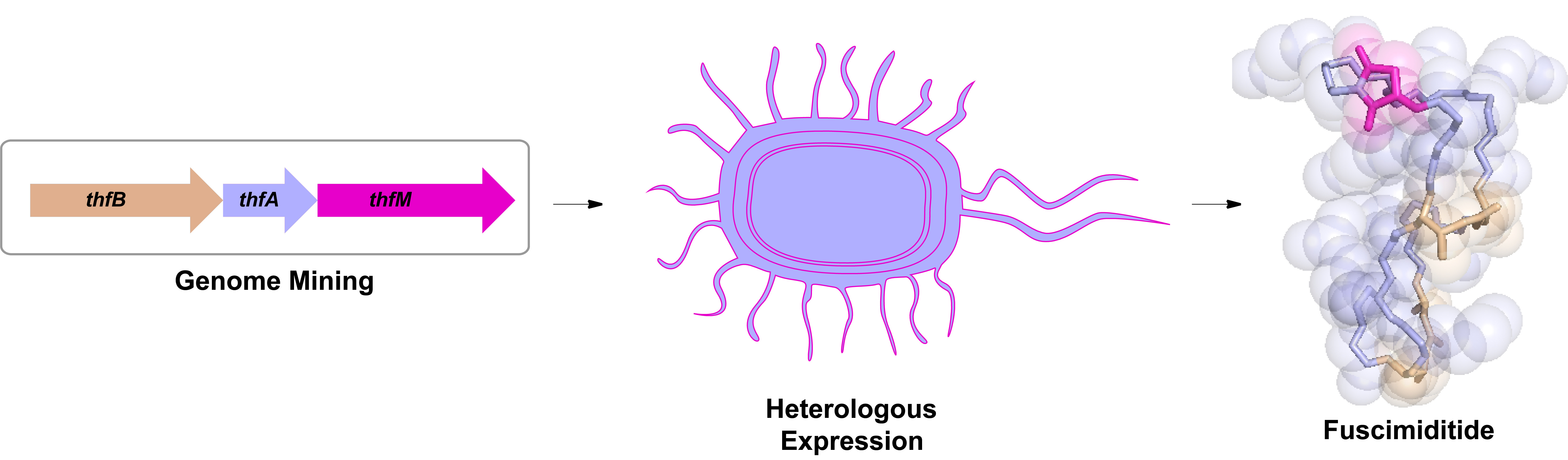
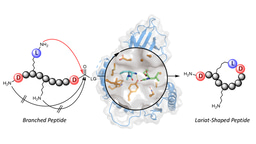
Figure 2. Workflow of fuscimiditide characterization. Genome mining revealed a graspetide-like cluster with a previously uncharacterized gene (thfM) in Thermobifida fusca. Subsequent cloning and heterologous expression in E. coli revealed three dehydrations catalyzed by ThfB and ThfM. Fully matured fuscimiditide is a stem-loop macrocyclic peptide connected via two ester crosslinks (wheat) with an aspartimide moiety (magenta) embedded within the loop region.
Over the course of 4 years, this work has become a labor of love, with my colleague Joe Koos laying the groundwork before I joined the Link lab in 2018. The project felt like a puzzle with every experiment bringing us closer to completing the picture of which amino acids were involved in the linkages, what the linkages were, and the role of each enzyme. There was even a point when I thought fuscimiditide was shaped like a pretzel! Although, I was later proven wrong by HMBC correlation studies which proved the structure to be a hairpin instead.
Let’s also not forget that there was a pandemic in the midst of this ongoing project. The uncertainty of the quarantine period and the supply chain shortages that followed really tested our resilience. We would like to acknowledge the work and support that the other Link lab members have put into making this paper possible, especially Li Cao for his insight into aspartimides through his own work on aspartimide containing lasso peptides [2], Brian Choi for his cloning expertise, Wai Ling Cheung-Lee for her NMR guidance, and Michelle Richardson and Heather White for their initial work on the project back in its early days.
- Lee, H., Choi, M., Park, J., Roh, H., Kim, S. Genome Mining Reveals High Topological Diversity of ω-Ester-Containing Peptides and Divergent Evolution of ATP-Grasp Macrocyclases. J. Am. Chem. Soc. 142, 3013-3023 (2020).
- Cao, L., Beiser, M., Koos, J.D., Orlova, M., Elashal, H.E., Schroder, H.V., Link, A.J.: Cellulonodin-2 and Lihuanodin: Lasso Peptides with an Aspartimide Post-Translational Modification. J. Am. Chem. Soc. 143, 11690-11702 (2021).
Follow the Topic
-
Nature Chemistry

A monthly journal dedicated to publishing high-quality papers that describe the most significant and cutting-edge research in all areas of chemistry, reflecting the traditional core subjects of analytical, inorganic, organic and physical chemistry.

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in