Bee-pollination promotes rapid divergent evolution in plants growing in different soils
Published in Ecology & Evolution and Plant Science

Evidence of divergence in plant traits between related plant taxa is usually attributed to a shift in pollinators and adaptation to different soil types. Such patterns arise when adaptation to one factor, such as soil type, leads to subsequent adaptation to the other factor, such as pollinator preference. This could happen because the distribution of pollinators covaries with soil types in a mosaic-like pattern, as proposed by the Grant-Stebbins model of pollinator-driven speciation, or because adaptation to soil reinforces differential attraction of pollinators upon secondary contact. Both scenarios involve changes in pollinator guilds as the main driver of the divergence in flower type and the subsequent emergence of pollination ecotype, though some studies suggest that pollination ecotypes can also arise as a by-product of habitat adaptation. Additionally, herbivory can play a significant role in this complex interplay by affecting floral aesthetic, inducing the synthesis of deterrent compounds in floral signals and increasing reward toxicity. As a result, herbivory constrains the evolution of floral traits by mediating conflicting selection pressures imposed by pollinators and herbivores on these traits. Therefore, understanding the intricate interplay between soil type, pollinators, and herbivores is essential for unraveling the complexities of macroevolutionary diversification in plants.
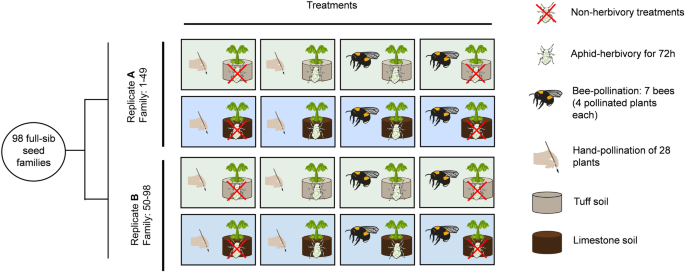
To address this issue, we conducted an experimental evolution study including 8 treatments with fast cycling Brassica rapa plants, encompassing a full factorial design of three different factors: soil (growing either in limestone or tuff soil), hand-pollination or bumblebee-pollination, with- or without aphid-herbivory. We studied the impact of soil, pollination, herbivory, and their interactions on driving divergent evolution in plant traits after 8 generations of selection. For herbivory, we infested plants at the two true leaves stage with 10 Brevicoryne brassicae aphids, and allowed aphids to feed and reproduce on plants for three days, after which aphids were removed. We used Bombus terrestris to induce pollinator mediated-selection, and limited visits to 7 bumblebees and 5 visits per bumblebees to ensure pollen limitation, providing a near-natural context for observing evolutionary response in plant traits. To control for the contribution of pollinator to plant evolution, we used hand-pollination, which represents evolution without pollinator-mediated selection.

Figure 1. Soil collection at the two sites location. (a) Dr. Luca Arrigo and Pr. Giovanni Scopece collecting limestone soil in Valico di Chiunze (40.719°N, 14.619°E). (b) Pr. Giovanni Scopece collecting tuff soil in Monte di Procida.
This ambitious experiment required massive labor and logistic work. Soil was collected in the region of Naples and involved the extraction of ca 500 kg of limestone and tuff soil, and its shipment to Switzerland. Soil collection required the help of many people who spent long hours during summer and autumn collecting soil (Figure 1). The experiment would have simply never started without these wonderful people who extracted and hand-sifted cube meters of soil. Upon reception of soil, the experiment spread across three years, starting in 2018 and ending in 2021. It entailed the cultivation of ca 17,000 plants and the use of ca 31,000 aphids for aphid-herbivory. Despite the challenges represented by aphid-herbivory, we successfully controlled aphid infestation manually throughout our entire experiment. Although this process was time-consuming, it proved to be the best approach, as tests prior to the experiment demonstrated that insecticides never completely eradicated aphids and often caused damage to plant tissues, particularly in plants growing in limestone soil. Therefore, we deemed preferable to not use pesticides, prioritizing a method that effectively managed aphids without side effects or soil contamination.

Figure 2. Differences in morphology between plants being visited (selected) by bumblebees (right side) versus those that have not been visited (left side), specifically within the context of plants evolving in tuff.
Throughout the experiment, bumblebee-mediated selection was strong, especially on traits such as plant height and flower number (see Figure 2). Irrespective of herbivory and soil, bee-pollinated plants experienced the stronger evolutionary changes, with bigger flowers, and fewer glucosinolates across generations. Interestingly, we found strong evidence of divergent evolutionary changes between bee-pollinated plants that evolved in our different soils. As evolution progressed, plants evolving in tuff soil and with bee-pollination grew taller, along with a significant shift in phenology, with a later flowering onset compared to its pairs in hand pollination and limestone treatments. With a longer vegetative stage, these plants produced a bigger leaf area, which allowed plants to stand taller by pollination time as well as sustain a higher flower production (Figure 3). Those contrasting evolutionary changes also involved other morphological traits such as nectar amount, or changes in individual scent and glucosinolate compounds. Additionally, we showed that aphid-herbivory, though it did not lead to higher defense levels, still compromised the evolution of flower attractiveness to pollinators. This reduced attractiveness in plants that evolved with herbivory likely resulted from diminished floral scent emission and reduced plant height (Figure 3c), both of which are key signals of attractiveness to pollinators in the here used species as well as many other plant-pollinator systems.

Figure 3. Impact of bee-pollination and aphid-herbivory on plant evolution. (a) Differences in morphology and phenology between treatments in early development. (b) Example of bee-first choices matrix including each plant line. In figures (a) and (b), different colors represent the different soil treatments (blue: limestone soil, green: tuff soil, black: generation one), while different shapes indicate different pollination treatments (circle: bee-pollination, squares: hand-pollination). (c) Differences in morphology in tuff line plants with bee-pollination when evolving without herbivory (left) or with herbivory (right). (d) Differences in morphology in limestone line plants with bee-pollination when evolving with herbivory (left) or without herbivory (right). (e) Differences in morphology in bee-pollination and non-herbivory line plants that evolved in tuff (left) or in limestone (right).
Our study provides experimental evidence that floral diversification can occur independently of shifts in pollinators. We demonstrate that the soil-pollinator effect serves as a mechanism leading to floral divergence without changes in pollinator composition. As the soil-pollinator effect drives population divergence in traits such as floral traits, we suggest that it may be followed by pollinator shifts. A shift in pollinators may reinforce divergence between populations as different pollinators differ in preferences and/or morphologies, thus aligning with the concept of the geographic mosaic of pollinator distribution.
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026





Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in