Cultivate rocks, catch Asgard archaea
Published in Microbiology

Asgard archaea and the eukaryotic origin. The emergence of a complex eukaryotic cellular plan is one of the greatest evolutionary marvels. We infer the past from the present. Present-day mitochondria carry genetic material that is clearly inherited from bacteria either belonging or related to the α-proteobacteria; present-day nuclei inherited archaeal genes responsible for core cellular functions executing the central dogma. It is thus now rather well accepted that an ancient union between an archaeon and a bacterium gave birth to a eukaryotic cell plan1. Figure 1 is a naïve depiction of this so-called eukaryogenesis model. However, the when, how, and who remain a mystery at large. The discovery of Asgard archaea has brought much excitement in the past few years as they were shown to be the closest archaeal relatives to the present-day eukaryotes2,3. Their images also sensationalized new speculations over how eukaryotic cellular features emerged4-6. Here is a brief account on the process that led to our recent discovery of new Asgard archaea species via in-lab cultivation of deep-sea rocks (Wu et al Nature Microbiol 2022).
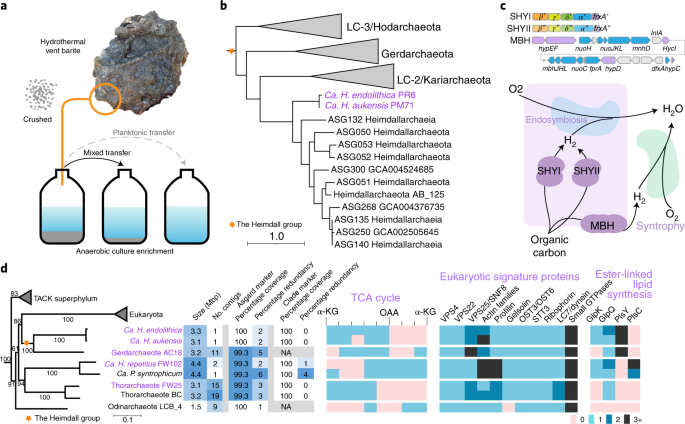
Cultivating rocks. The fall of 2019 was the first time I saw a definitive presence of the Heimdall group (or the original Heimdallarchaeota), the branch of Asgard archaea that was shown in some analyses to be the closest to eukaryotes, in the rocks. [Note that the definition of many phyla, including Heimdallarchaeota, is undergoing rearrangement due to the progressive expansion of our understanding of Archaea.] It had been about a year since my colleague Antoine Crémière and I had crushed a part of a fist-sized rock into grains of sand, soaked it in anaerobic artificial sea water, bottled it up, and placed it in a 40°C incubator. It was no ordinary rock, though. It was retrieved near Diane’s vent [named in memory of marine scientist Diane Poehls Adams] of the Auka vent field in the Southern Pescadero basin, the deepest known hydrothermal vent system in the Pacific Ocean, by my colleague Daan Speth during a research expedition in the fall of 2017. We found that these rocks host diverse communities of microbes and wanted to understand how they function in the local ecosystem [see Daan’s article on the global analyses of the microbial diversity in the nearby sediment7].
A stunning image of the Diane’s vent featured on the cover of the February 2022 issue was taken in 2018 with the remotely operated vehicle SuBastian during cruise number FK181031, aboard Schmidt Ocean Institute's research vessel Falkor, led by co-chief scientists Dave Caress, Robert Zierenberg and our own Victoria Orphan. Having a ‘habitat’ of comfort at the lab bench and only tagging along occasionally with the team out to sea, I could not have imagined working with samples from such an unlikely place if it were not owing to the Orphan lab’s rich seafaring experience and the many willing colleagues. Since a post-lunch coffee with Daan and Antoine in the Orphan lab social room that kicked off the project, we gained help from many skillful and kind colleagues experimentally (Igor Antoshechkin, Aditi Narayanan, and Stephanie Connon) and computationally (Alon Philosof and Roman Barco), even when they scrambled to combat all kinds of curve balls thrown by the COVID19.
Emergence of a Heimdallarchaeum clone from ‘nothing’. Antoine and I had been growing rocks for a while but had always been trying to maintain the native community using methane (unpublished work), the most abundant local energy source and a strong greenhouse gas whose fate the Orphan lab has been disentangling for nearly 20 years. Meanwhile, I had grown curious about the thousands of microbial lineages that live on and in the rocks but do not directly consume methane. A cascade of interactions must exist among these microbes as well as between them and their environment, such as the rocks. Applying nutrient perturbations is one of the approaches we took. By adding different energy source other than methane, the microbial community became very quickly displaced, and the apparently insignificant or even undetectable ones became dominant. The expansion from ‘nothing’ to something in a microbial community brought an unexpected advantage. Metagenomic sequencing often face the inherent complication where related species cross-interfere, leading to fragmentation and chimeras during bioinformatic assembly, whereas a ‘clean’ clone that is drastically different from the others is in principle much less prone to these issues. A Heimdallarchaeota archaeon, which we later named Candidatus Heimdallarchaeum endolithica, emerged in such a fashion and, to our knowledge, became the first of its clade to be recovered a circularized complete genome. Several other complete and contiguous genomes from our other incubations followed owing to such an approach. They include the circular genome of Ca. Heimdallarchaeum aukensis from an incubation derived from the above 2018 expedition co-led by Victoria.
Unveiling mobile (genetic) elements. Once the first Ca. Heimdallarchaeum genome was circularized, Daan and I started poking at it from various directions and observed two ‘weird’ features. One is the existence of giant genes that came in pairs, each encoding proteins 5000 amino acids. The other was an unusual peak when we mapped the sequencing reads onto the genome. We had suspected that the former was a misannotation, and the latter a misassembly, but later both were found to be Heimdallarchaeal mobile elements, most likely integrative viruses. This is owing to the capture of the full suite of CRISPR/Cas systems within the genomes, which contain repeat sequences that would have been broken up into fragments in metagenomic assemblies without recent advances in Nanopore long-read sequencing. I named the former ‘Aloposon’, after the twin giants Aloedae in Greek mythology, and the existence of 4 such elements within two genomes eased the initial concerns of misannotations. The latter appeared to replicate bidirectionally in a circular form, indicated by the beautiful V-shaped read-mapping pattern and confirmed via PCR. Mobile elements are agents that shuffle genes across species, and we found that they inherited various nucleic acid-processing enzymes from bacteria and bacteriophages, indicating a complex evolutionary entanglement between Asgard archaea and bacteria.
Horizontal gene transfer, the pan-Asgard gene repertoire, and the eukaryotic genome expansion. It has been found that the eukaryotic genomes garnered their gene content from both bacterial and archaeal domains8,9, and curiously, the fraction of bacteria-derived genes increases with genome size. Our estimate suggests a similar trend between Asgard archaea and eukaryotes both in terms of gene numbers and functional division. In the paper, we also discussed the striking difference between the scaling properties of the ‘bacterial’ faction and ‘archaeal’ fraction in these genomes. We do not have a clue regarding the mechanism that led to such a difference, a topic very much open to future explorations. However, the observed parallelism suggests that understanding how Asgard archaea (or archaea in general) regulate genome size and the gene content may provide a clue for the early stages of gene acquisition that established the eukaryotic genomes.
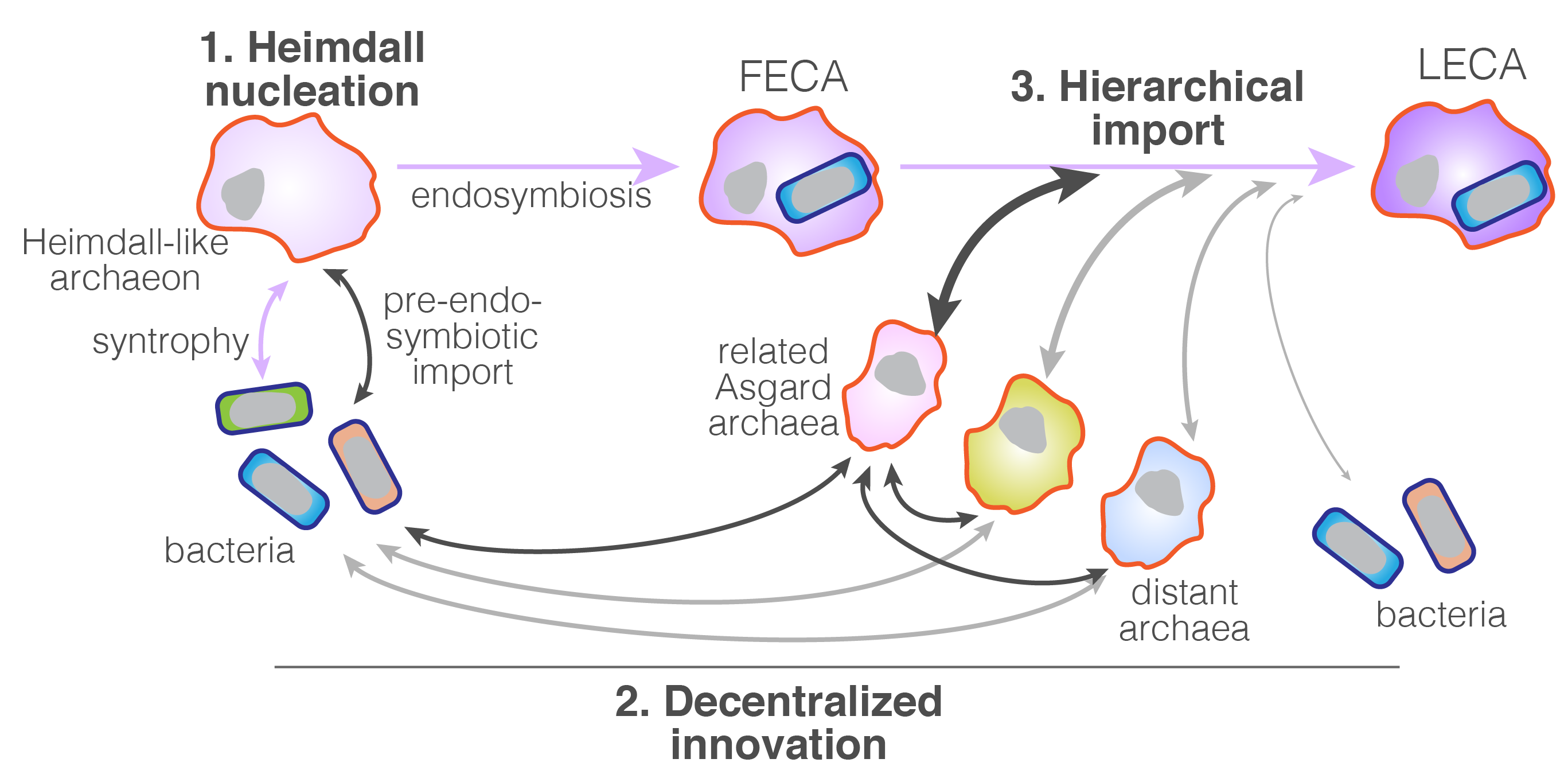
One of the great disparities between eukaryotes and prokaryotes is their overall genome sizes range. While Asgard archaea are shown to be the closest to eukaryotes, their genome architecture is still very much archaeal. Their sizes range from ~1.5 Mb to ~ 4.5Mb, seemingly unremarkable when compared with eukaryotes, which ranges from less than 10Mb up to over 100 Gb (count in picograms!10). The smallest eukaryotic genomes are mostly thought to be a result of genome reduction due to their symbiotic or parasitic lifestyle. Under the constraint of small genomes, prokaryotes make it up by pertaining a ‘pangenome’, where closely related species together maintain a large repertoire of genes that they can shuffle relatively painlessly to allow rapid adaptation11. We postulate that such a context within the Asgard archaea is critical for the early stage of eukaryotic evolution. A line of evidence for it is the fact that beyond the core genes that almost all major Asgard archaea lineages share with eukaryotes, there are many eukaryotic features that are lineage specific. To sum it up, we put forth a conceptual framework called the Heimdall nucleation - Decentralized innovation - Hierarchical import model (Fig. 2) to account for the gene content expansion that led to the emergence of eukaryotic gene repertoire.
1 Eme, L., Spang, A., Lombard, J., Stairs, C. W. & Ettema, T. J. G. Archaea and the origin of eukaryotes. Nature Reviews Microbiology 15, 711-723, doi:10.1038/nrmicro.2017.133 (2017).
2 Spang, A. et al. Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature 521, 173-179, doi:10.1038/nature14447 (2015).
3 Zaremba-Niedzwiedzka, K. et al. Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature541, 353-358, doi:10.1038/nature21031 (2017).
4 Imachi, H. et al. Isolation of an archaeon at the prokaryote–eukaryote interface. Nature 577, 519-525, doi:10.1038/s41586-019-1916-6 (2020).
5 Avcı, B. et al. Spatial separation of ribosomes and DNA in Asgard archaeal cells. The ISME Journal 16, 606-610, doi:10.1038/s41396-021-01098-3 (2022).
6 Salcher, M. M. et al. Visualization of Lokiarchaeia and Heimdallarchaeia (Asgardarchaeota) by Fluorescence In Situ Hybridization and Catalyzed Reporter Deposition (CARD-FISH). mSphere 5, e00686-00620, doi:doi:10.1128/mSphere.00686-20 (2020).
7 Speth, D. R. et al. Microbial community of recently discovered Auka vent field sheds light on vent biogeography and evolutionary history of thermophily. bioRxiv, 2021.2008.2002.454472, doi:10.1101/2021.08.02.454472 (2021).
8 Alvarez-Ponce, D., Lopez, P., Bapteste, E. & McInerney, J. O. Gene similarity networks provide tools for understanding eukaryote origins and evolution. Proceedings of the National Academy of Sciences 110, E1594, doi:10.1073/pnas.1211371110 (2013).
9 Koonin, E. V. & Yutin, N. The Dispersed Archaeal Eukaryome and the Complex Archaeal Ancestor of Eukaryotes. Cold Spring Harbor Perspectives in Biology 6 (2014).
10 Pellicer, J., Fay, M. F. & Leitch, I. J. The largest eukaryotic genome of them all? Botanical Journal of the Linnean Society 164, 10-15, doi:10.1111/j.1095-8339.2010.01072.x (2010).
11 McInerney, J. O., McNally, A. & O'Connell, M. J. Why prokaryotes have pangenomes. Nature Microbiology 2, 17040, doi:10.1038/nmicrobiol.2017.40 (2017).
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in