Dying to organise
Published in Materials, Physics, and Cell & Molecular Biology
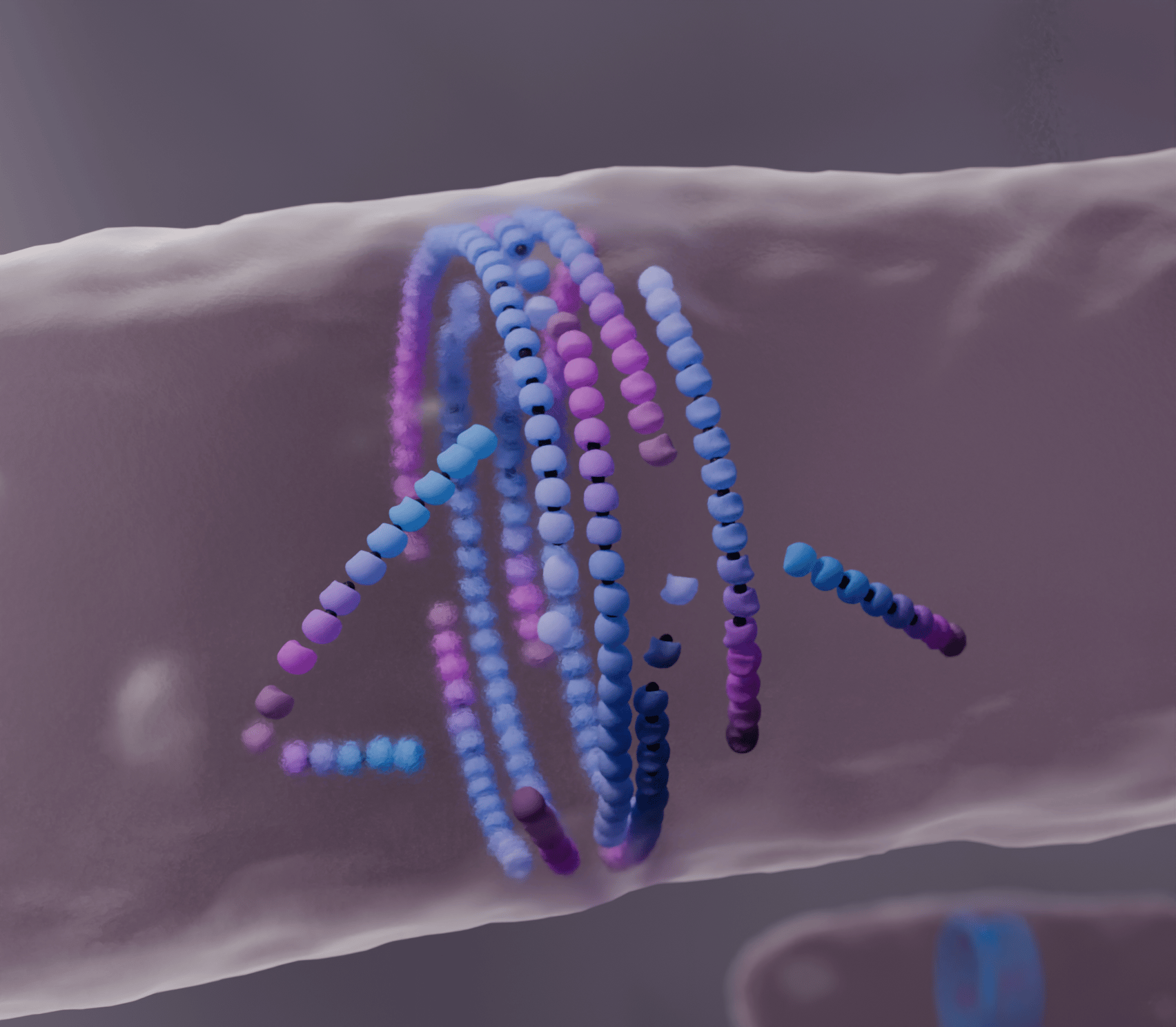
Death and birth in cytoskeleton organisation
Cells rely on the cytoskeleton, a dynamic collection of interlinking filaments, to maintain their shape and internal organisation. The cytoskeleton constantly (re)organises itself to mediate essential cellular functions like cell division or movement. This dynamic cytoskeletal organisation requires energy consumption: chemical energy in the cell is transformed into functional work of the cytoskeleton. Here, the energy is typically consumed in two ways: i) filaments can consume energy themselves to drive their own dynamic growing and shrinking, called turnover and ii) associated motor proteins can consume energy to slide filaments past each other. The latter scenario has been extensively studied, while the first case remains largely underexplored despite constituting an essential aspect of cytoskeletal organisation.
In particular, prokaryotic cells, like bacteria and archaea, are lacking molecular motors capable of generating active pushing or pulling forces to drive motion of filaments. However, by undergoing constant turnover, their cytoskeleton is still capable of functional collective self-organisation across scales. Here we focus on bacterial FtsZ filaments – dynamic filaments that assemble into a ring around the midcell of a bacterium and drive its division. These filaments consume energy to grow on one end while shrinking on the other, causing them to appear motile even though individual proteins remain static. Inspired by FtsZ treadmilling and the bacterial division ring, in this work we explore how turnover of filaments mediates their collective functional self-organisation.
1,2,3, align! – 1 question, 2 countries, 3 teams
Our lab is fascinated by how inanimate molecular objects come together inside the cell to create life. Using computer simulations, we strive to disentangle the mechanisms by which molecules come together to give life-like functions. A fascinating example of such a process is cell division, which across the tree of life, is driven by self-organisation of cytoskeletal proteins into a filamentous ring around the cell body. We initially wanted to understand whether the division ring in bacteria can generate a constriction force that would partially squish the cell and had decided to model its self-organisation from individual protein molecules. However, we were unable to assemble the ring by pure filament growth – we would always end up in a disordered state that did not resemble a ring. We then went back to the literature and realised that filaments in cells are not just growing – they are constantly growing from the head and shrinking from the tail by consuming chemical energy, which is called treadmilling. It initially seemed odd to us that the cell would spend this energy just to constantly turnover its filaments, but then again, without it we were unable to assemble a self-organised ring. So, a clear question emerged for us: Does turnover help filaments to collectively self-organise?
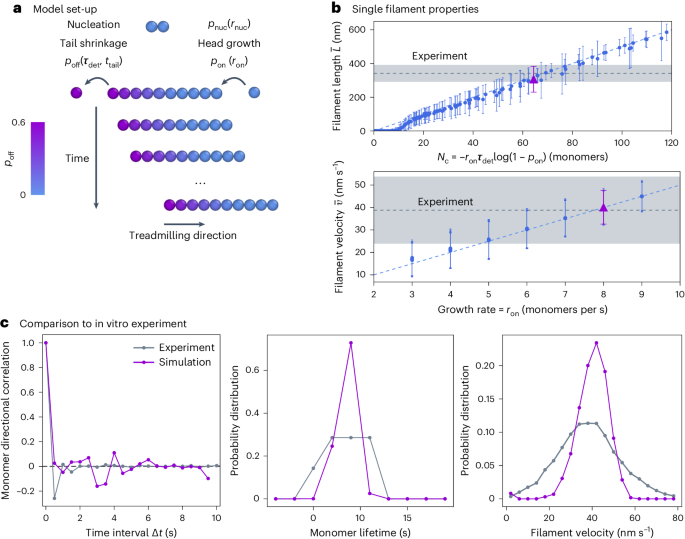
We thus set off to design an appropriate minimal coarse-grained model that would incorporate filament growth and shrinkage and enable the simulation of many filaments together, both on a generic flat surface and then on a curved surface such as that of a bacterial cell.

In the spirit of looking for minimal models and general physical principles, we described protein filaments as beads connected by springs. However, coupling filament growth-shrinkage kinetics to diffusive dynamics of the polymers in simulations proved to be complex. It took some time and computational effort to develop a model of filaments that grow from the head at an imposed rate and depolymerise from the tail in a time-dependent manner, capturing the characteristic aging of treadmilling polymers. With a working model in hand, we were then able to study how treadmilling affects filament organisation. We found that, while passive, purely diffusive filaments remain disordered over time, treadmilling alone drives large-scale ordering of the filaments. Our model shows that this active death and birth of filaments enables the dissolution of misaligned components leading to collective alignment, and that disabling this activity results in disordered systems. So dying and growing leads to ordering!
It was around that time that we attended an online Physics meets Biology conference, where we met Séamus Holden and his team, who were working on imaging of bacterial ring formation in live cells. They had just collected beautiful experimental data supporting precisely the idea that the birth and death kinetics was essential for the formation of the ring. Fuelled by this chance synchronicity, we joined forces and used our model to make sense of these experimental findings. Working together enabled us to make sure that our physical model is in a biologically-relevant regime, and to test the model on live cell data.

Satisfyingly, we found that FtsZ rings in our simulations behaved in the same way as the Bacillus subtilis division rings that Holden’s team imaged: rings formed very quickly but took long to grow and mature. Our model was able to make sense of the ring dynamics observed in their data, demonstrating that it is the mortality of these filaments that drives ring formation and maturation.
Shortly after, our group moved from London to Vienna, setting up at the Institute of Science and Technology Austria. Coincidentally, at the same Institute the group of Martin Loose had been working on the in vitro reconstitution of FtsZ filaments. They happened to have collected amazing data on FtsZ collective organisation in vitro on flat membranes. This serendipitous encounter allowed us to quantitatively test our model predictions for dissolution-driven alignment against a clean and controlled in vitro system, which we found matched extremely well our simulations.
Thinking back to how this project came together, it is curiosity and the right interdisciplinary platforms that brought the right people together. Exploring ideas beyond our comfort zones, disciplines and geographic borders, and discussing science openly made the discovery possible. The main result of this project might just be the collaborative relationships and new research avenues built along the way.
A key takeaway: dying drives alignment
By undergoing turnover, treadmilling filaments like FtsZ form dynamic large-scale structures that spontaneously and robustly align while remaining efficiently sensitive to external biases. Monomers in the filaments age by consuming energy and eventually depolymerise from the tail. This monomers loss is usually balanced by filament growth at the head, but if polymerisation is disrupted filaments tend dissolve and die. Typically, misaligned filaments tend to have more random collisions with their neighbours than their aligned counterparts, which results in a greater disruption to their growth. Consequently, misaligned treadmilling filaments tend to die quickly which, over time, results in the overall selection of collectively aligned filaments. Filament mortality thus allows for collective organisation by mediating the dissolution of misaligned components in the system.

Illustration: Claudia Flandoli
So what? Conclusions and implications
Follow the Topic
-
Nature Physics
This journal publishes papers of the highest quality and significance in all areas of physics, pure and applied.


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in
On the study: “Death and Birth in Cytoskeleton Organisation via FtsZ Treadmilling”
✦ Visionary Perspective: Integrating AI into Cytoskeletal Self-Organisation StudiesThe current work presents an elegant minimal physical model to explain how local dynamics—filament birth and death—can drive large-scale structural organisation in prokaryotic cells. However, I believe that the integration of Artificial Intelligence (AI) and Machine Learning (ML) approaches could greatly enrich both the analytical power and predictive potential of this research. I propose the following new directions:
1. Deep Learning for Live Imaging Analysis❖ Novel Idea: Apply deep learning models (e.g., convolutional neural networks or recurrent networks) to analyze live-cell imaging datasets of FtsZ filament dynamics in real time.
Automatically detect ring formation or failure.
Quantify the rate and spatial distribution of filament turnover.
Predict filament ageing and collective alignment transitions.
Energy-state color gradients (blue to purple) can be mapped as time-series inputs for the AI model to recognize filament “life cycle” patterns.
2. Reinforcement Learning for Filament Behavior Optimization❖ Bold Proposal: Build a reinforcement learning (RL) environment where intelligent agents simulate the behavior of individual filaments.
Each agent controls its own polymerization/depolymerization rate.
The system is trained to evolve toward optimal ring formation or minimal energy expenditure.
Emergent behavior may reveal unforeseen self-organization strategies.
This could lead to the concept of a “smart filament” that learns to self-organize collectively under minimal rules and local feedback.
3. Unsupervised Learning to Detect Anomalous Dynamics❖ Research Opportunity: Apply unsupervised machine learning (e.g., autoencoders, clustering algorithms) to detect unusual or inefficient organizational patterns in filament simulations or experimental data.
-
-
-
4. Graph Neural Networks (GNNs) for Filament Interaction ModelingIdentify early signs of disordered growth.
Classify organization regimes (ordered, metastable, chaotic).
Guide biological experiments toward unexplored configurations.
Given that FtsZ filaments form interactive networks across the membrane surface, Graph Neural Networks (GNNs) offer a promising framework:
-
-
-
5. Inverse Simulation via AI for System ReconstructionModel filaments as nodes with dynamic edge weights (e.g., contact, alignment).
Capture how local interactions influence global ring architecture.
Simulate complex, large-scale organisation in a computationally efficient way.
❖ Ambitious Direction: Use inverse-design machine learning models to reconstruct initial conditions from observed outcomes.
Provide the AI model with a final ring structure; it returns the most likely set of parameters (growth rates, number of filaments, cell curvature) that could produce such a configuration.
This approach would significantly accelerate hypothesis generation in cytoskeletal biophysics.
✦ Concluding Vision: AI as a Catalyst for Discovering Self-Organising PrinciplesIntegrating AI into cytoskeletal studies enables a shift from descriptive observation to predictive modeling and ultimately design-driven biology.
This will not only deepen our understanding of fundamental self-organisation but could also:
Aid in designing synthetic cells or programmable biological architectures.
Offer insights into failure mechanisms in pathological division (e.g., in cancer).
Bridge molecular dynamics and emergent cell-scale phenomena.