Expanding inovirus diversity: the rise of the small and inconspicuous
Published in Microbiology
## Read the full paper at: https://rdcu.be/bLije ##
One may first ask: why even look for inoviruses ? The short answer is that inoviruses are bacteriophages with two fascinating properties: (i) their short (~5-15kb) single-stranded DNA genome is enclosed in an unusual filamentous particle, and (ii) they replicate through a unique “chronic” cycle by which they can produce new infectious virions without killing their host cell (1). Until now, inoviruses have mainly been studied for their biotechnological potential e.g. in phage display (2) or for their impact on the infectivity and toxicity of major human pathogens including Vibrio Cholerae, Neisseria meningitidis, and Pseudomonas Aeruginosa (3).
 compared with typical dsDNA bacteriophages (B, C, & D) observed with Transmission Electron Microscopy (TEM).")
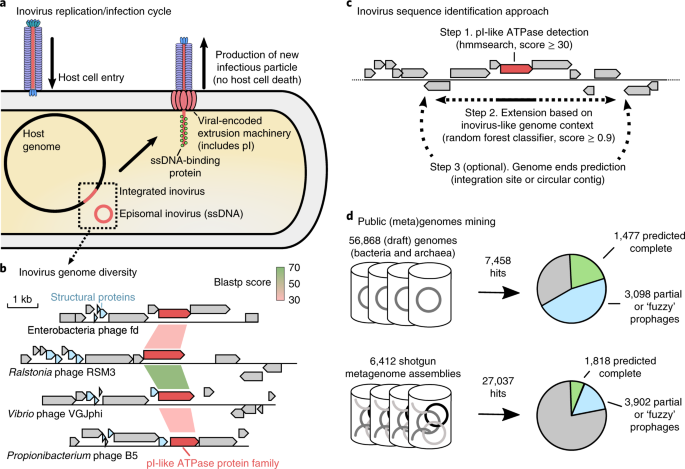
Back in 2012, the answer to Mart’s question was “No, we don’t really see any inoviruses”. Fast-forward to 2017 and we still have only 56 inovirus in the genome databases, and virtually none detected in metagenomes. At this point, we decided to start over and design a new tool to specifically detect these inoviruses, leveraging a combination of custom marker genes and machine learning approaches (4). Once we were confident we could robustly identify inoviruses, we mined a large set of > 50,000 genomes and > 5,000 metagenomes and, finally, here they were: 10,995 inovirus sequences, representing an estimated 5,964 species.

Some of these
detections fell in the “hard-to-believe” or
“something-must-be-wrong” category: four inoviruses had been
presumably identified in archaeal genomes. This was surprising
because inoviruses are known to infect bacteria, and historically
viruses infecting bacteria and archaea have been thought to be
entirely distinct, although this strict evolutionary border is
progressively getting blurred as we explore more of the virosphere
(5). Still, to convince ourselves that these were real, we worked
with Rebecca Daly from the Wrighton Lab at CSU, who grew these
archaea hosts in the lab and performed induction experiments, i.e.
she attempted to get the virus to replicate and form new particles.
Using PCR, we could confirm the presence in the sample of a circular,
excised form of the complete predicted inovirus genome. Now, the next
step will be to find conditions leading to a high replication rate
for this virus so that we can (hopefully) observe newly-formed
filamentous viral particles using TEM, and further understand the
biology of these first archaea-infecting inoviruses.

Another puzzling observation was the extensive diversity of genes encoded by inoviruses, which rivaled the one found across phage groups with much larger genomes. One of our initial hypothesis was that some of these inovirus “hypothetical proteins” were here to shut down the CRISPR-Cas system of their host. We selected the best candidates for this function, partnered with the JGI Synthetic Biology team to get the genes synthesized, and sent these constructs to Adair Borges in the Bondy-Denomy Lab at UCSF to test their function. However, instead of CRISPR-interacting proteins, what Adair discovered was evidence of superinfection exclusion: some inovirus-encoded proteins provide additional defense to their host cell by preventing other (larger) viruses to infect the same cell. This suggests inoviruses could increase their host’s fitness by providing additional defense against (unrelated) phages, and reinforce the need for further investigation of virus-virus interactions (6).
So coming back to Mart’s initial question, what did we learn when trying to look for inoviruses ? First, inoviruses are much more diverse and likely more important than previously thought, in many hosts and across virtually every biome. Second, the fact that we could identify so many virus sequences that had gone (somewhat) unnoticed is a stark reminder of how incredibly diverse the viral world is, and that we have not yet reached the end of our “virosphere exploration” journey (by a long shot). And finally, this project also exemplifies the amazing opportunities now available when leveraging interdisciplinary teams, especially when establishing a “feedback loop” between “dry” data analysis and “wet” laboratory bench experiments, each guiding the other as hypotheses are refined and patterns emerge.
For more information, see: Roux et al., Cryptic inoviruses revealed as pervasive in bacteria and archaea across Earth’s biomes, Nature Microbiology, doi: https://doi.org/10.1038/s41564-019-0510-x / https://jgi.doe.gov/learning-to-look-inoviruses/. Data generated in this project are available at https://genome.jgi.doe.gov/portal/Inovirus/Inovirus.home.html.
(1) Mai-Prochnow, A. et al. ‘Big things in small packages: The genetics of filamentous phage and effects on fitness of their host’. FEMS Microbiol. Rev. 39, 465–487 (2015). doi: 10.1093/femsre/fuu007
(2) Recently recognized by a Chemistry Nobel Prize to George Smith and Gregory Winter: see the greatvisual summary by C&EN here
(3) Sweere, J. M. et al. Bacteriophage trigger anti-viral immunity and prevent clearance of bacterial infection. Science. 363, eaat9691 (2019). doi: 10.1126/science.aat9691
(4) Machine learning spots treasure trove of elusive viruses, Amy Maxmen, Nature News. (2018) https://www.nature.com/articles/d41586-018-03358-3. doi: 10.1038/d41586-018-03358-3
(5) Prangishvili, D., Bamford, D. H., Forterre, P. & Iranzo, J. The enigmatic archaeal virosphere. Nat. Rev. Microbiol. 15, 724–739 (2017). doi: 10.1038/nrmicro.2017.125
(6) Díaz-Muñoz, S. L., Sanjuán, R. & West, S. Sociovirology: Conflict, Cooperation, and Communication among Viruses. Cell Host Microbe 22, 437–441 (2017). doi: 10.1016/j.chom.2017.09.012
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.





Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in