Microbes contribute to nitrogen and arsenic cycling even in sediments of the deepest ocean trench on Earth (Mariana Trench)
Published in Microbiology

Hadal trenches are the deepest regions in the world’s oceans located between 6000 m and 11,000 m (20,000 to 36,000 ft) below sea level. These deep-sea trenches, named after the Greek god of the underworld, Hades, are geological formations that were created in the Cenozoic/Cretaceous period (65.5/100 million years ago) due to subduction of one of Earth’s tectonic plates under another (Jamieson et al., 2010). The collision of one oceanic crustal plate against another can result in the downward bending of ocean crust along the edges of each plate, forming a deep trench, and ultimately, the burial, or subduction of one of the two plates underneath the other. Hadal trenches are the most remote marine habitats, and much like Hades, are known for their extreme characteristics, including high hydrostatic pressures (> 1,000 atm), absence of light and nutrient scarcity that collectively, create intense/hostile conditions for life. Despite these unfriendly conditions and the geographical isolation of hadal seabeds and trench bottoms, they nonetheless are turning out to host dynamic and complex microbial communities (Glud et al., 2013) that exhibit possibly high degrees of endemism (taxa/species found uniquely in these habitats) (Jamieson et al., 2011).
The 11-kilometer deep Mariana Trench is the deepest of the 27 hadal trenches in our global ocean, and Challenger Deep, which is located at the south end of the Mariana Trench, is the deepest location along the trench, and therefore, the deepest recorded oceanic habitat on Earth. Challenger Deep has a V-shape topography which is characteristic of most trenches. This funnel-type topography drastically influences the amount of detrital organic material and associated nutrients that accumulate along the trench slope vs. along the trench bottom axis. These precious sources of carbon and energy fuel the microbial communities in benthic habitats, including in hadal trenches. The origin of these sources of particulate organic and inorganic nutrients is from regional seismic/volcanic activities as well as associated slumping of sediments into the trench, and from biological production in the overlaying water column (for example, dead and sinking organisms and/or their exudates, and faecal matter). Due to the technical challenges associated with sampling sediments and waters at these extreme hadal depths, little is still known about life in hadal trench sediments and bottom waters. As the deepest hadal trench on Earth, Challenger Deep microbial communities and their metabolic capabilities are the least explored.
In our study we collected samples of sediment cores across the slope and bottom-axis of the Challenger Deep (locations ranging from ~6 to 11-km depth) to study in situ hadal microbial communities and to provide insights into their metabolic activities (Figure 1).
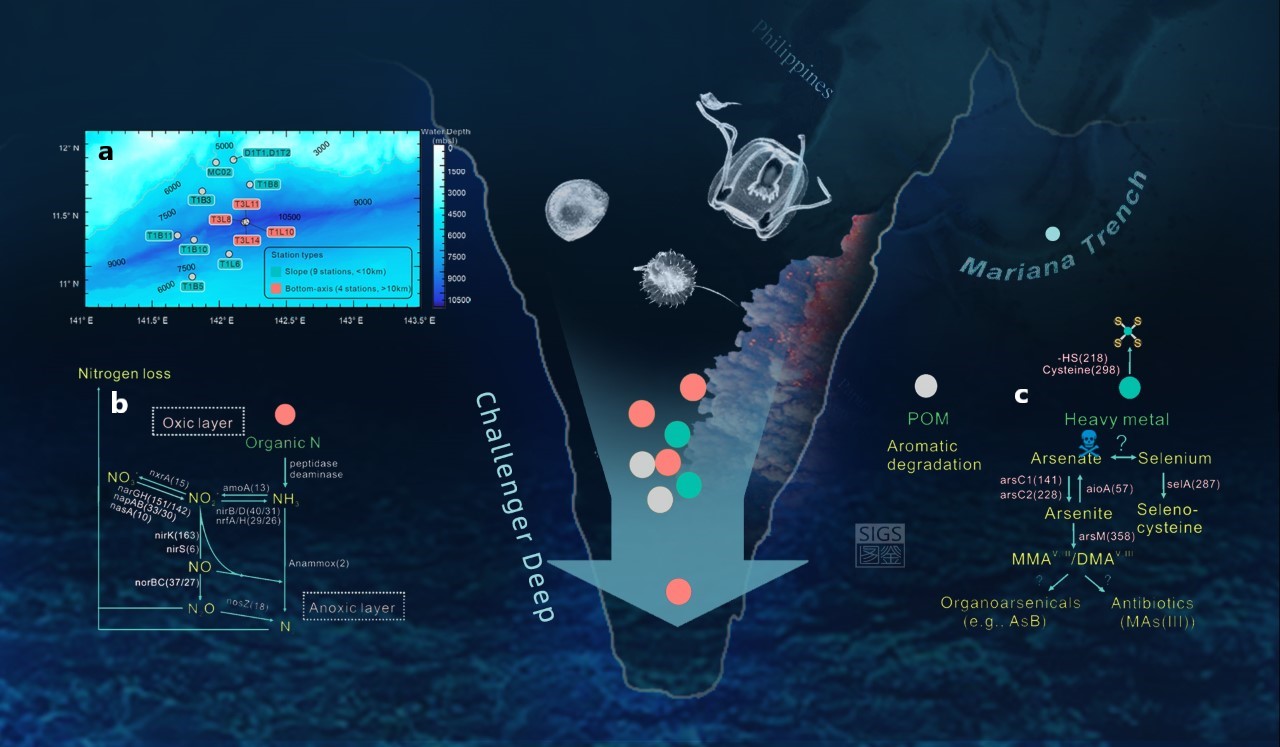
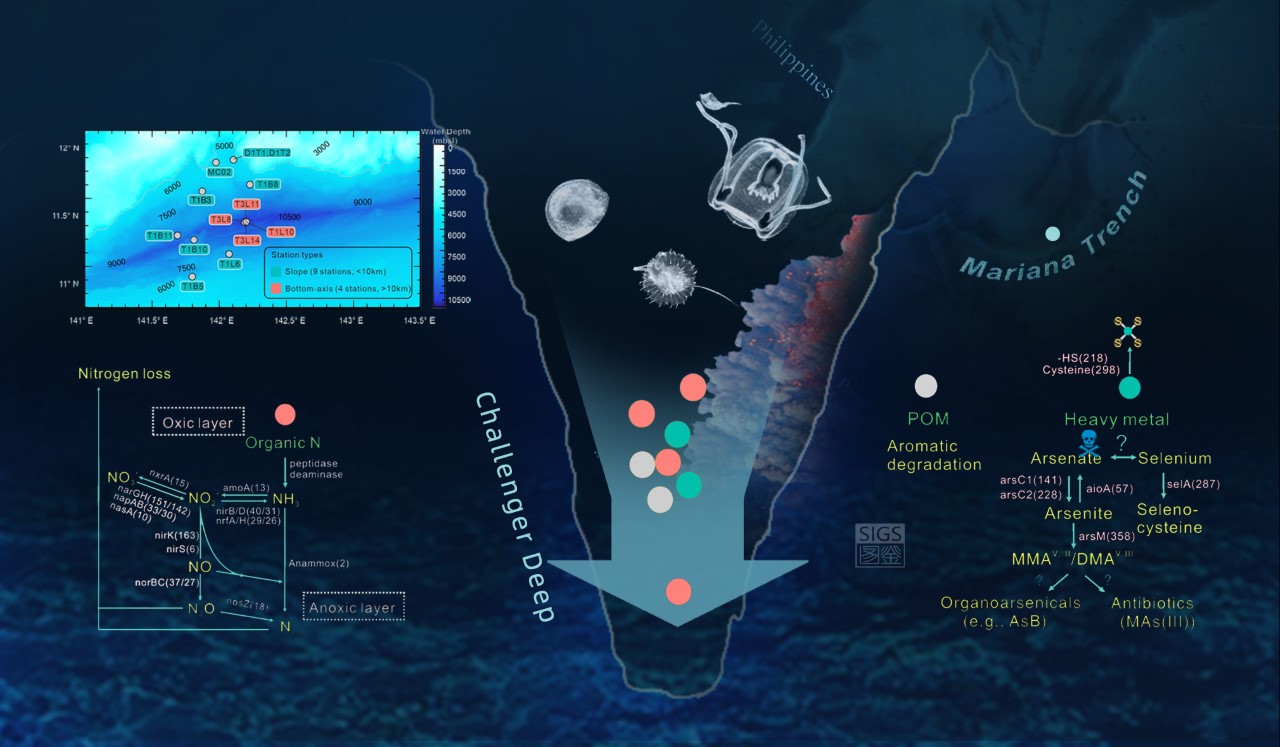
Figure 1: Overview of sampling sites and potential microbial metabolic activities in Challenger Deep (CD) sediments. Sampling sites (a), suggested nitrogen cycling and nitrogen release (b) and microbial biotransformations of arsenic and selenium (c) in CD sediments.
We analyzed 586 metagenome-assembled genomes (partially-assembled genomes of bacteria and archaea recovered within the pool of environmental DNA) and we found that the composition of the prokaryotic (bacteria/archaea) community is distinct between the bottom axis and slope sites of Challenger Deep. Of the recovered small subunit ribosomal RNA gene signatures, we identified 26% that were novel (defined as <97% identity to references from publicly available prokaryotic databases), with novelty increasing with water and core sediment depths. By examining recovered gene transcripts (genes transcribed into RNA and recovered within the total RNA pool) and the assembled prokaryotic genomes we determined that the microbial communities identified in Challenger Deep are very likely utilizing ammonia and nitrate for gain energy (instead of oxygen which is almost absent at the surface of sediment cores, and disappears below the sediment surface; Figure 1b).
The usage of ammonia and nitrate by hadal microbes has been previously suggested for Challenger Deep sediments (e.g., Nunoura et al., 2018). However, these important earlier observations were based on marker genes (key genes that describe only one step of the biochemical process of interest), and not on genomic data that could support presence of the complete pathways for energy gain (e.g., ammonia oxidation and denitrification). The nitrogen in ammonia and nitrate is released after respiration into the overlying water column as nitrogen gas. Nonetheless, the variable availability of nutrients and the variable composition of in situ microbial communities appears to create a heterogeneous (uneven) distribution of microbiota with different metabolisms, and this heterogeneity will shape the release of nitrogen from different sites, as well as transformations of diverse organic and inorganic compounds in this hadal realm.
Other metabolic activities identified include the ability of Challenger Deep microbes to recycle proteins for energy, and to utilize simple and complex hydrocarbons as carbon sources. Saturated hydrocarbons (e.g., n-alkanes, which are carbon atoms arranged in a single chain) are shown to accumulate in surficial sediments at 11 km- depth in Challenger Deep, and likely derive from sinking particles and/or synthesized in situ from heterotrophic communities which are distinct from other known hydrocarbon-producing marine organisms (Liu et al., 2019).
Our study also showed that the identified hadal microbes have the potential to biotransform heavy (e.g., arsenic) and trace minerals (e.g., selenium). Some of these metabolisms may resemble ancient types of metabolism used for energy gain (Figure 1c). Alternatively, they may be endemic metabolic traits that enhance survival (e.g., production of arsenic-based antibiotics, osmoregulation, aminoacid analogues) in this hostile environment of Challenger Deep.
For details, please see our manuscript https://www.nature.com/articles/s41467-022-29144-4.
1. Jamieson, A.J., Fujii, T., Mayor, D.J., Solan, M., Priede, I.G (2010). Hadal trenches: the ecology of the deepest places on Earth. Trends Ecol Evol. 25(3):190-7. doi: 10.1016/j.tree.2009.09.009.
2. Glud, R. N., Wenzhöfer, F., Middelboe, M., Oguri, K., Turnewitsch, R., Canfield, D. E., Kitazato, H. (2013). High rates of microbial carbon turnover in sediments in the deepest oceanic trench on Earth, Nat. Geosci., 6: 284–288. https://doi.org/10.1038/ngeo1773.
3. Jamieson, A.J. (2011). Ecology of Deep Oceans: Hadal Trenches. In eLS, (Ed.). https://doi.org/10.1002/9780470015902.a0023606.
4. Nunoura, T., Nishizawa, M., Hirai, M., Shimamura, S., Harnvoravongchai, P., Koide, O., Morono, Y., Fukui, T., Inagaki, F., Miyazaki, J., Takaki, Y., Takai, K. (2018). Microbial diversity in sediments from the bottom of the Challenger Deep, the Mariana Trench. Microbes Environ. 33(2):186-194. doi: 10.1264/jsme2.ME17194.
5. Liu, J., Zheng, Y., Lin, H, Wang, X., Li, M., Liu, Y., Yu, M., Zhao, M., Pedentchouk, N., Lea-Smith, D-J., Todd, J.D., Magill, C.R., Zhang, W-J., Zhou, S., Song, D., Zhong, H., Xin, H., Yu, M., Tian, J., Zhang, X-H (2019). Proliferation of hydrocarbon-degrading microbes at the bottom of the Mariana Trench. Microbiome 7:47. https://doi.org/10.1186/s40168-019-0652-3.
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Biosensing
Publishing Model: Hybrid
Deadline: Sep 30, 2026


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in