
Forest diversity mediated by pathogens
I was introduced to the Janzen-Connell hypothesis and negative conspecific density dependence (CDD) around 2015, when I was working on my Master’s degree. It was fascinating that pathogens, by definition harmful organisms, could actually generate and sustain community level plant diversity. Specialized (or relatively specialized) pathogens could create a zone of repulsion under adult trees, reducing the recruitment of their offspring, and leaving space for other species. This negative ‘conspecific density dependence’ (CDD), prevents any one species from dominating and allows a diversity of species to establish. I was working with a professor and her PhD student to see whether this applied within species – are species more successful when grown under genetically more dissimilar individuals of the same species? Because I was excited by all things mycorrhizal fungi, I was hired as a work study student to score all the seedling roots for arbuscular mycorrhizal fungi (the mycorrhizal relationship 70-80% of plant species worldwide depend on). That means I mounted many roots on slides, and quantified colonization of different mycorrhizal structures under the microscope. Even back then, I wondered if these beneficial fungi might ‘pull’ seedlings closer to their adults, and mitigate the enemy driven mechanism that had been studied and/or implied as the dominant driver of CDD. We did find within species microbial specialization (see our paper); however, in the end, we did not find any strong effect of mycorrhizal fungi within our focal species.
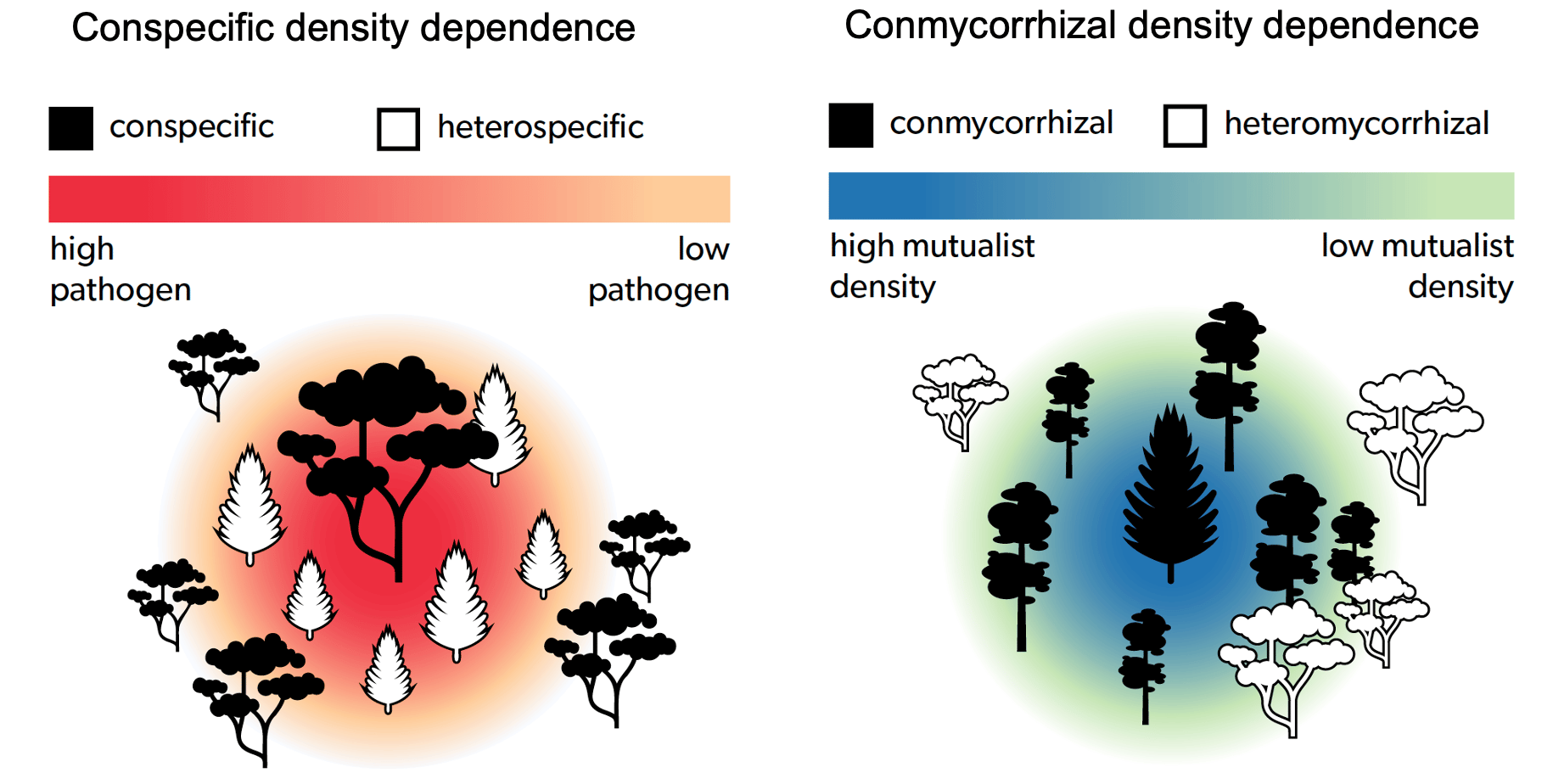
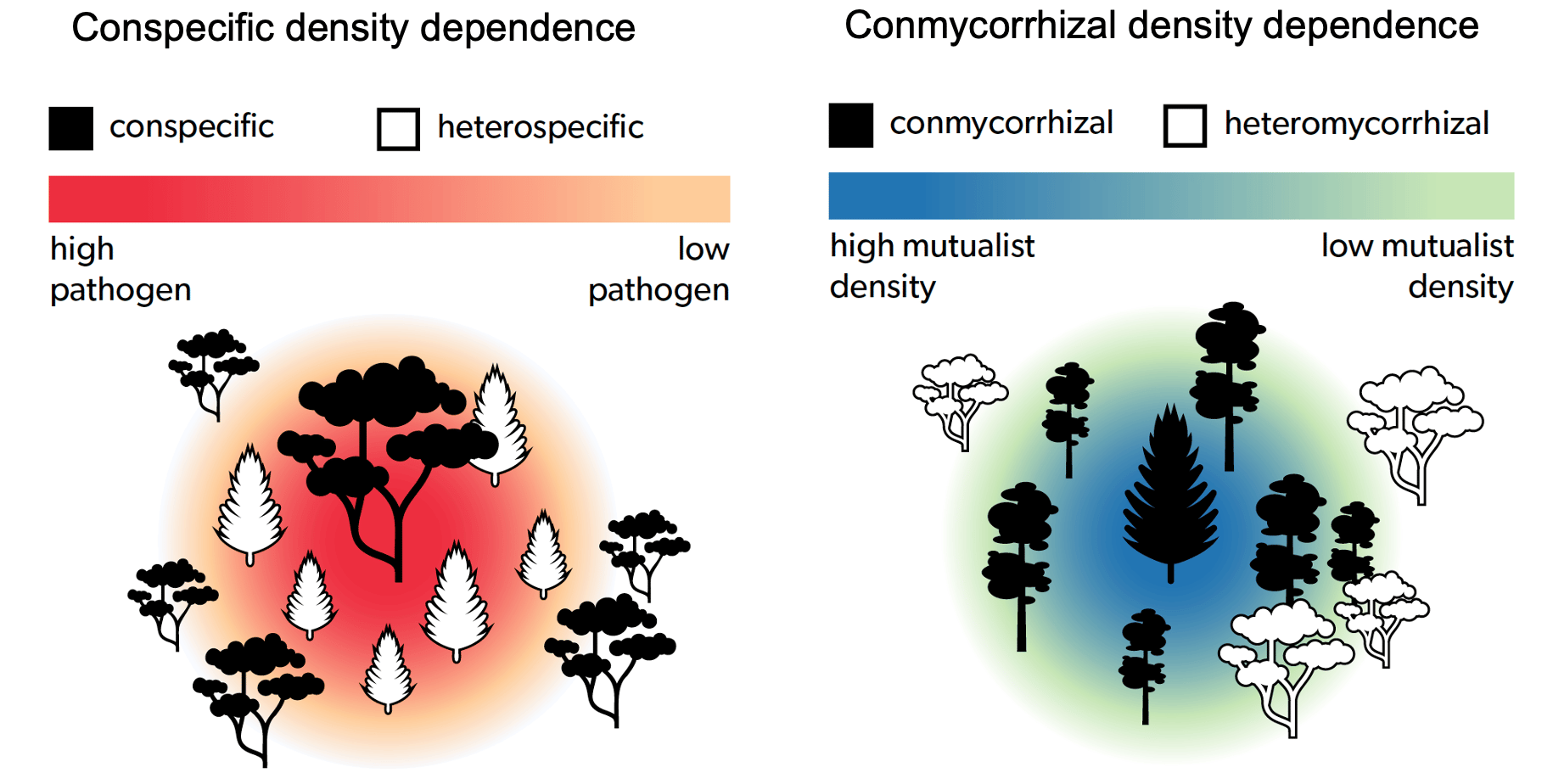
Conceptual figures depicting negative conspecific density dependence (CDD) in recruitment due to hypothesized higher densities of species-specific enemies around a conspecific adult tree and positive conmycorrhizal density dependence (CMDD) in recruitment due to hypothesized shared mutualists with an adult tree of a conmycorrhizal heterospecific species.
A rejected proposal
I went through the next six years without explicitly revisiting this hypothesis – until I applied for my postdoctoral fellowship. I wanted to leverage the global ForestGEO network to test whether these mycorrhizal fungi might matter. How do mutualists and pathogens interact to influence CDD? Importantly, might be there be a fundamental difference in CDD between plants associated with the two major mycorrhizal fungal types? Previous work showed arbuscular mycorrhizal (AM) species have stronger CDD than ectomycorrhizal (EM) plants, potentially due to differences in specificity and pathogen protection (EM species are thought to be higher in both). But, this work was confined to regional comparisons and investigated this at a broader scale using only tree data (i.e. overlooking the microbiome itself). We wanted to do this at the global scale, and directly study the pathogen and mycorrhizal components of the microbiome. I reached out to Joe LaManna, since I had read his paper many times in writing my proposal. He spent hours going through his code and models with me. I worked with researchers at ETH Zurich in the Crowther Lab to write my postdoctoral fellowship application. I submitted the proposal and got an interview. The proposal was rejected. But, we began building contacts and setting up the infrastructure to do the project.
An accepted proposal!
I began contacting every plot PI, with the help from Joe and good 'old fashioned' cold emailing. In the meantime, I worked on my second proposal to carry out this work. With some preliminary data and the buy in of around 30 plots, I submitted an application for the Swiss National Science Foundations’ Postdoctoral Fellowship. On a bus coming back from holiday, I opened my email and saw a message from the Swiss National Science Foundation - I had gotten the two year grant to complete the full project! The funding covered a tree level comparison, and a follow up study directly sampling soils, characterizing microbial communities, and linking this to CDD and diversity. Our first step was testing for a AM versus EM signature of CDD in trees.
Mycorrhizal types mediate CDD at the global scale
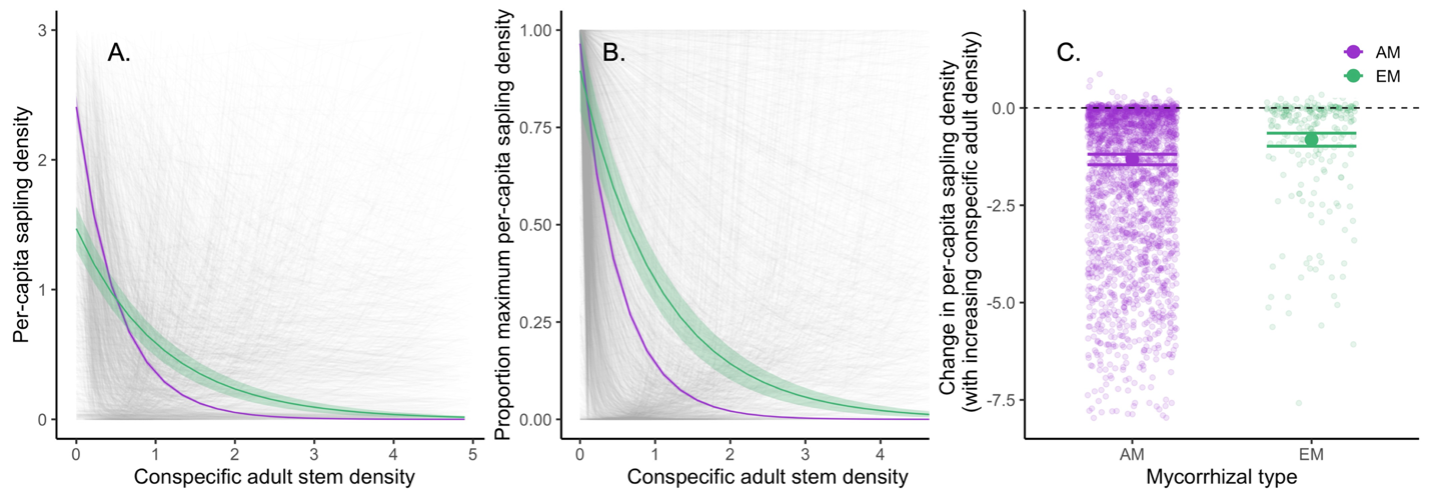
In this study, we used census data across 43 large-scale forest inventory plots worldwide, including nearly 3 million stems and over 4,000 species. Ultimately, we found that EM tree species show weaker negative CDD, likely due to greater host specificity or pathogen protection compared to tree species associating with arbuscular mycorrhizal fungi. This, coupled with the well-known global latitudinal gradient in mycorrhizal types – with ectomycorrhizal plants increasing with absolute latitude – suggests forest mycorrhizal strategy is an essential component of the overall weakening of CDD and lower tree diversity observed at higher latitudes. We also showed that both mycorrhizal types exhibit positive conmycorrhizal feedbacks, with trees benefitting from the presence of heterospecific neighbors that form the same mycorrhizal type, potentially by tapping into shared neighboring mycorrhizal fungi. This positive mycorrhizal feedback at the community level may explain why many forest stands exhibit mycorrhizal bimodality globally, with stands where both mycorrhizal strategies coexist occurring far less than expected by chance. Collectively, these findings suggest that mycorrhizal interactions may play a foundational role in global forest diversity patterns and structure. These findings bring to light the important role of the forest fungal microbiome, mutualisms, and positive feedback in maintaining global patterns of forest biodiversity.
What’s next?
We are working through soil samples from around 30 ForestGEO plots to directly link the microbial community composition to changes in CDD and global diversity patterns (see this blog I co-wrote about our sampling on Barro Colorado Island, Panama). We are working through around 1300 soil samples from across the world to identify what microbes are present (sequencing) and soil chemical properties. Ultimately, the two main goal are to (1) understand the biogeographical drivers of mycorrhizal fungi and pathogens at our plots and (2) see whether CDD and plant diversity are directly related to microbial diversity and composition.
Follow the Topic
-
Communications Biology
An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.

Related Collections
With Collections, you can get published faster and increase your visibility.
Artificial Intelligence Methodology in Structural Biology
Publishing Model: Hybrid
Deadline: Nov 30, 2026
Healthy Aging
Publishing Model: Open Access
Deadline: Dec 31, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in
Congrats Camille, very interesting work emphasising the need to look at microbiome more intensively than usually do when we are dealing with ecosystem interactions and resilience.