Potential therapeutic strategy for recurrent ovarian cancer with a novel NAMPT inhibitor
Published in Cancer
Background
Advanced ovarian cancer (OV) is a deadly disease. OV, which is the most lethal gynecological malignancy, is often diagnosed at late stages [1]. Due to the difficulty of complete removal of the tumor in advanced stage, multidisciplinary treatment combining surgery and chemotherapy with a platinum-based drug regimen is recommended [2]. While platinum-based chemotherapy is effective in the majority of OV patients, over 80% of advanced-stage cancers eventually relapse due to chemo-resistance, necessitating the development of new treatment strategies [3].
NAD+ is an essential co-enzyme involved with metabolic processes required for survival and growth. NAD+ is synthesized from three different pathways, including the Preiss–Handler pathway, generating NAD+ from nicotinic acid (NA) via nicotinic acid phosphoribosyltransferase (NAPRT); the de novo synthesis pathway, generating NAD+ from tryptophan (Trp) via quinolinate phosphoribosyl transferase (QPRT); and the salvage pathway, generating NAD+ from nicotinamide (NAM) via Nicotinamide phosphoribosyltransferase (NAMPT) [4]. Cells rely on the salvage pathway as the main source of NAD+ [5]. Several NAMPT inhibitors such as FK-866, GNE-617, GNE-618, CHS-828 have shown antitumor effects in a variety of cancers [6]. KPT-9274 is a first-in-class, orally bioavailable NAMPT inhibitor designed to provide energy depletion, DNA repair inhibition, cell cycle arrest and growth inhibition. It targets two enzymes, NAMPT and p21 activated kinase 4 (PAK4) [7]. The expression level of PAK4 is often elevated in various types of cancers at DNA, RNA, or protein level, and is proposed as a diagnostic biomarker for cancer [7]. While its therapeutic efficacy has been demonstrated preclinically in several cancer types [8, 9, 10], the potential of KPT-9274 in OV remains unexplored.
In this study, we investigated these mechanisms using platinum-resistant 3D-cultured spheroids, a preclinical model which morphologically mimics the tumor mass in the fluid that circulates in the patient’s abdominal cavity (ascites) and thus strongly reflect the characteristics of recurrent OV [11].
Key findings
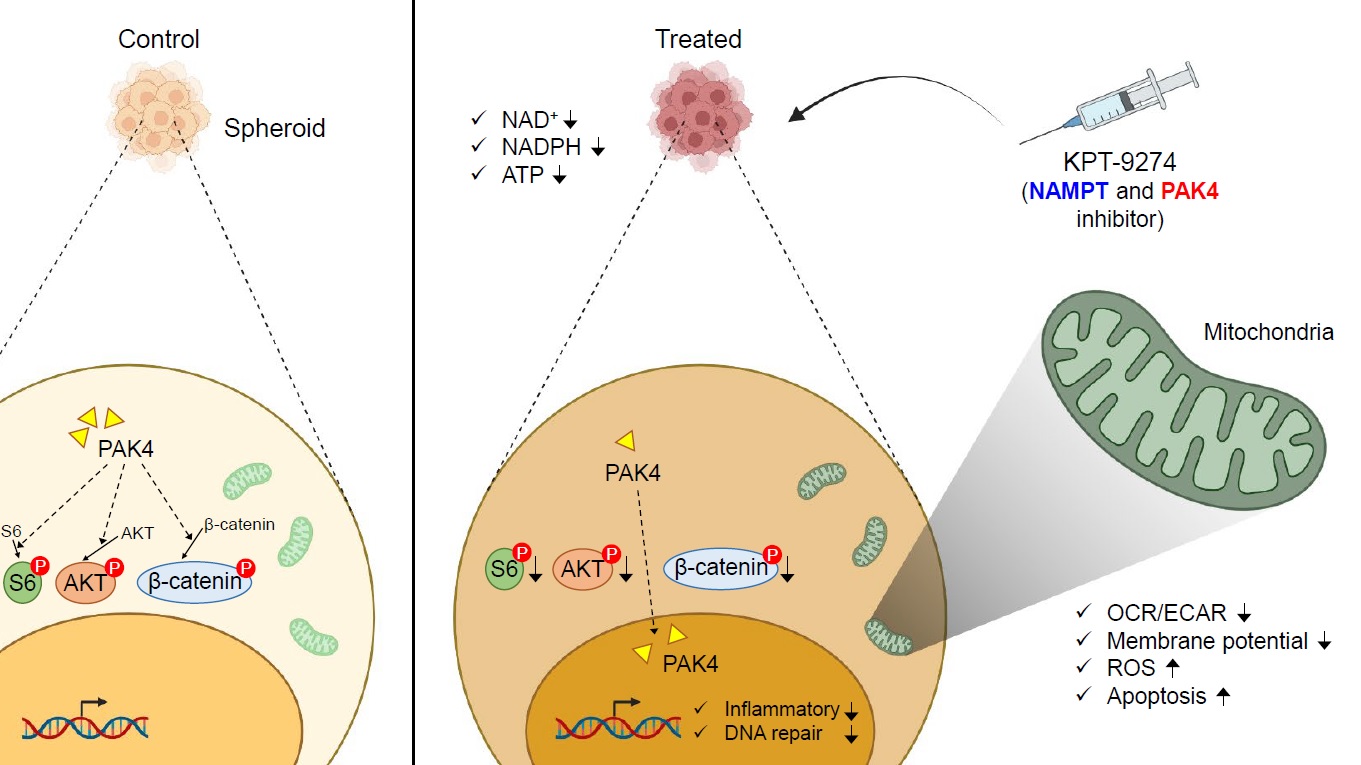
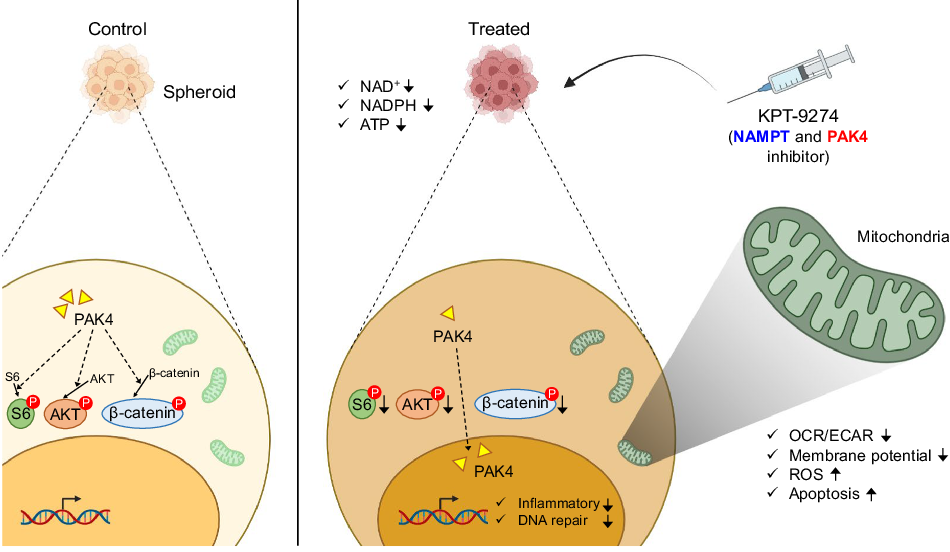
Our first step was to investigate the sensitivity of OV cell lines to KPT-9274 using the 3D-cultured spheroids preclinical model. Interestingly, we found that KPT-9274 not only inhibited the cell proliferation, but also suppressed NAD+, NADPH, and ATP production in NAMPT-dependent cell lines with platinum-resistance, indicating that KPT-9274 could be a promising treatment for OV patients who have developed resistance to platinum-based therapies. Given that KPT-9274 suppressed ATP production, we hypothesized that KPT-9274 decreased mitochondrial function. Indeed, we observed that KPT-9274 down-regulated established measures of mitochondrial function, including oxygen consumption rate with Seahorse experiments [12], Tetramethylrhodamine Methyl Ester Perchlorate (TMRM) [13], while upregulating markers reflecting mitochondrial damage, such as MitoSOXTM Red [14] and cleaved caspase 3/7 [14]. These results suggest that KPT-9274 suppresses mitochondrial function.
To further characterize the impact of NAMPT and PAK4 in OV, we next compared patients with high and low NAMPT expression using RNA-sequencing data from The Cancer Genome Atlas (TCGA). Notably, Gene Set Enrichment Analysis revealed that patients with high NAMPT expression exhibited enriched gene sets related to inflammation in OV patients. Similarly, we compared patients with high and low PAK4 expression. Among high PAK4 patients, pathways related to gene repair and cell proliferation (e.g. G2M checkpoint, mTORC1 signaling, Wnt/β-Catenin signaling, and PI3K-AKT-MTOR signaling) were found to be up-regulated. These results highlight the impact of high NAMPT and high PAK4 in OV patients. Additionally, to corroborate the results obtained from TCGA datasets, the impact of KPT-9274 treatment in the preclinical model was evaluated through RNA sequencing. As anticipated, we demonstrated that KPT-9274 regulates cell proliferation by suppressing gene expression and pathways related to inflammatory signaling, DNA repair, and tumor growth, consistent with findings from public datasets.
Finally, considering that PAK4 regulates the phosphorylation of various proteins related to cell proliferation, including RAPTOR, S6, AKT, and β-catenin [15, 16, 17], we evaluated how KPT-9274 affects their phosphorylation. PAK4, which was predominantly localized in the cytoplasm before KPT-9274 treatment, migrated into the nucleus after treatment. In parallel with the shift of the localization of PAK4, the cytoplasmic expression levels of RAPTOR, Phospho-S6 Ribosomal Protein (Ser235/236), Phospho-AKT (Ser473), and Phospho-β-Catenin (Ser675) were decreased. Overall, these findings suggest that KPT-9274 impedes cell proliferation by reducing cytoplasmic kinase activity through altering PAK4 localization.
Take home message
Drug treatment for recurrent OV faces severe challenges due to platinum-chemotherapy resistance. Here, we provide a strategy to overcome this hurdle by dual inhibition of NAMPT and PAK4 with KPT-9274. The inhibitor suppresses not only NAD+, but also NADPH, mitochondrial function, gene expression related to inflammatory signaling and DNA repair, and phosphorylation of several proteins associated with cell proliferation (RAPTOR, S6, AKT, and β-catenin) in NAMPT-dependent cell lines exhibiting platinum-resistance. These findings emphasize the importance of targeting the drug to patients whose tumor growth depends on NAMPT for the effective use of KPT-9274 in clinical. We believe that NAMPT inhibitors, including KPT-9274, could soon lead to successful clinical translation for biomarker-selected, platinum-resistant recurrent OV.
References
- Torre LA, Trabert B, DeSantis CE, Miller KD, Samimi G, Runowicz CD, et al. Ovarian cancer statistics, 2018. CA Cancer J Clin. 2018;68(4):284-96.
- Armstrong DK, Alvarez RD, Bakkum-Gamez JN, Barroilhet L, Behbakht K, Berchuck A, et al. Ovarian Cancer, Version 2.2020, NCCN Clinical Practice Guidelines in Oncology. J Natl Compr Canc Netw. 2021;19(2):191-226.
- Lheureux S, Gourley C, Vergote I, Oza AM. Epithelial ovarian cancer. Lancet. 2019;393(10177):1240-53.
- Lucena-Cacace A, Umeda M, Navas LE, Carnero A. NAMPT as a Dedifferentiation-Inducer Gene: NAD(+) as Core Axis for Glioma Cancer Stem-Like Cells Maintenance. Front Oncol. 2019;9:292.
- Nacarelli T, Fukumoto T, Zundell JA, Fatkhutdinov N, Jean S, Cadungog MG, et al. NAMPT inhibition suppresses cancer stem-like cells associated with therapy-induced senescence in ovarian cancer. Cancer Research. 2019:canres.2830.019.
- Wei Y, Xiang H, Zhang W. Review of various NAMPT inhibitors for the treatment of cancer. Front Pharmacol. 2022;13:970553.
- Rane CK, Minden A. P21 activated kinase signaling in cancer. Semin Cancer Biol. 2019;54:40-9.
- Subedi A, Liu Q, Ayyathan DM, Sharon D, Cathelin S, Hosseini M, et al. Nicotinamide phosphoribosyltransferase inhibitors selectively induce apoptosis of AML stem cells by disrupting lipid homeostasis. Cell Stem Cell. 2021;28(10):1851-67 e8.
- Mogol AN, Zuo Q, Yoo JY, Kaminsky AZ, Imir OB, Landesman Y, et al. NAD+ Metabolism Generates a Metabolic Vulnerability in Endocrine-Resistant Metastatic Breast Tumors in Females. Endocrinology. 2023;164(6).
- Qasim SL, Sierra L, Shuck R, Kurenbekova L, Patel TD, Rajapakshe K, et al. p21-activated kinases as viable therapeutic targets for the treatment of high-risk Ewing sarcoma. Oncogene. 2021;40(6):1176-90.
- Gheytanchi E, Naseri M, Karimi-Busheri F, Atyabi F, Mirsharif ES, Bozorgmehr M, et al. Morphological and molecular characteristics of spheroid formation in HT-29 and Caco-2 colorectal cancer cell lines. Cancer Cell Int. 2021;21(1):204.
- Muller B, Lewis N, Adeniyi T, Leese HJ, Brison DR, Sturmey RG. Application of extracellular flux analysis for determining mitochondrial function in mammalian oocytes and early embryos. Sci Rep. 2019;9(1):16778.
- Creed S, McKenzie M. Measurement of Mitochondrial Membrane Potential with the Fluorescent Dye Tetramethylrhodamine Methyl Ester (TMRM). Methods Mol Biol. 2019:1928:69-76.
- Alam SR, Wallrabe H, Christopher KG, Siller KH, Periasamy A. Characterization of mitochondrial dysfunction due to laser damage by 2-photon FLIM microscopy. Sci Rep. 2022;12(1):11938.
- Li Y, Shao Y, Tong Y, Shen T, Zhang J, Li Y, et al. Nucleo-cytoplasmic shuttling of PAK4 modulates beta-catenin intracellular translocation and signaling. Biochim Biophys Acta. 2012;1823(2):465-75.
- Han C. Dan, Ricardo J. Antonia, Albert S. Baldwin. PI3K/Akt promotes feedforward mTORC2 activation through IKKα. Oncotarget. 2016; 7(16): 21064–21075.
- Cheng F, Li M, Thorne RF, Liu G, Zhang Y, Wu M, et al. P21-Activated Kinase 4 Pak4 Maintains Embryonic Stem Cell Pluripotency via Akt Activation. Stem Cells. 2022;40(10):892-905.
Follow the Topic
-
Cancer Gene Therapy
The essential gene and cellular therapy resource for cancer researchers and clinicians, keeping readers up to date with the latest developments in gene and cellular therapies for cancer.



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in