The DNA Loss Model (DNA-LM), more than 20 years contributing to explain neuropeptide evolution
Published in Ecology & Evolution, Protocols & Methods, and Anatomy & Physiology
In 2002, while I was doing my Ph.D. in Biological Sciences at the Faculty of Sciences of the National Autonomous University of Mexico, together with my directors Drs: Jesús Valdes, Samuel Zinker and Hugo Aréchiga. We postulated an evolutionary model based on DNA loss, which generated neuropeptide precursors of two phylums, which possibly corresponded to the molecular ancestor of the molluscan APGWamide (APGW), the Red Pigment Concentrating Hormone (RPCH) of crustaceans and the Adipokinetic Hormone (AKH) of insects.
All started by trying to obtain a possible organization of the RPCH gene of the crabs Procambarus clarkii and Cherax quadricarinatus by designing a strategy to quantify its messenger RNA (mRNA) and validate its participation in the circadian cycle of crustacean retinal pigments and cuticle. We began by reporting in 2002 that the RPCH gene of the crab Callinectes sapidus had an intron in the same position as the AKHs gene of Schistocerca nitens.
Simultaneously, we established the relevance of the results of Christensen et al. (1978, 1979), who demonstrated the relevance of the amino acids at the carboxyl end of the RPCH (QLNFSPGWG) to concentrate pigments and their similarity to the APGW of mollusks reported by Kuroki et al. (1990) and to the AKH II (QLNFSTGWG) of Schistocerca gregaria and Locusta migratoria reported by Siegert et al. (1985).
Thus, with the then scarce information from the APGW precursor mRNAs of a snail, a mussel and the California sea hare (Lymnaea stagnalis, Mytilus edulis, Aplysia californica), the RPCH of a pair of crabs (Carcinus maenas, C. sapidus) and of the AKHs of grasshoppers (L. migratoria, S. gregaria, S. nitans), cockroach (Blaberus discoidalis), hornworm (Manduca sexta) and fruit fly (Drosophila melanogaster), we demonstrated that the change in nucleotide alignment parameters of each precursor showed that the codons for the first four amino acids of the RPCHs and AKHs were located in two regions of the APGW precursors that do not produce neuropeptides; whereas the last four amino acids were homologous to one of the APGW copies.
Based on the above, we hypothesized that the conserved codons between the APGWs, RPCH and AKH were the product of natural selection to maintain their physiological function by binding to their receptor, whereas the non-homologous regions were unique to the APGW precursors and that these were possibly lost during evolution, if only, there was a duplication of the gene. Therefore, in the new gene, the conserved codons of RPCH/AKH were retained and lost codons unique to APGW. With this view, by joining and translating the conserved codons of the APGWs with respect to those of the RPCHs and AKHs, neuropeptides homologous to the RPCH or AKH were generated and named as virtual peptides because they were generated in silico.
In addition, to determine the possible loss of codons of the new gene, we consider that, if this event had occurred, the conservation or loss of nucleotides in the position of the introns in the gene should be observed. That is, both the original and the new gene should have introns in the same position. We demonstrated this when we determined that the nucleotides in the cDNA of the APGW precursors showed homology with the codons where the intron of the RPCH gene and the second intron of the AKHs were located. This suggested that the precursor gene for APGW would have an intron in that region reflecting the possible ancestral gene (Martínez-Pérez et al. 2002).
Sometime later, in 2007, we continued with the validation of the model by analyzing the binding of all the conserved codon domains in the APGW precursors of the three mollusks with respect to the precursors of the RPCHs and AKHs. The result was surprising, as precursors containing one to three copies of APGW and one copy of a neuropeptide similar to RPCH or AKH were obtained, which were named Neuropeptide Precursor Predictive (NPP). This result confirmed the possibility that the genome of one species had the gene for APGW and the other with the codons of an NPP with codons for molluscan APGW and arthropod RPCH or AKH. That is, the NPP would generate two neuropeptides with different activities, which at that time was unthinkable or inconceivable.
In searching for this species, we determined in silico that the APGW, RPCH, AKH and virtual RPCH/AKH peptides showed homology with copies of the precursor LWamide (LW), which was first described in the genus Hydra (DNGPMTGLWG) (Leitz et al. 1994, 1995), with the only difference being that the leucine was between the glycine and tryptophan of APGW and the last four amino acids of RPCH and other AKH (GPMTGLWG), which further supported the possibility of the existence of a species with an NPP gene for APGW/RPCH-AKH (Martínez-Pérez et al. 2007). Thus, we hypothesized that the gene could be in genomes of species whose body plan was similar to species of the Cambrian Explosion, for example, the Atlantic horseshoe crab Limulus polyphemus, the Pacific mole crab Emerita analoga or species of the class Polyplacophora, however, we were unable to obtain conclusive data due to the limitations of experimental methods at the time.
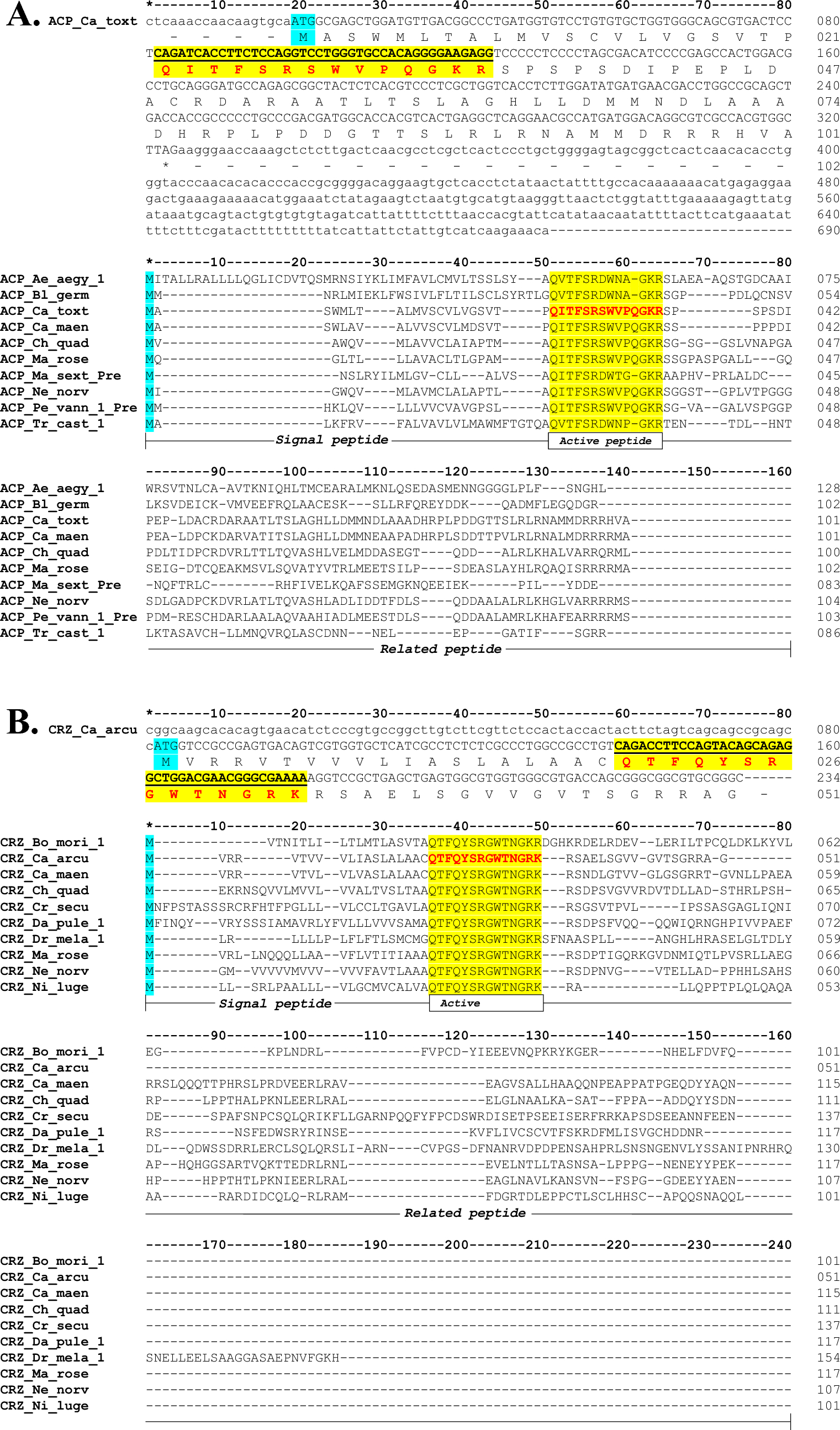
However, sometime later, with the development of next-generation sequencing (NGS), other research groups began to characterize invertebrate genomes and transcriptomes. In 2010, the genome of A. californica was published (Fiedler et al. 2010) and we were able to confirm that the positions of the introns in the APGW gene proposed in DNA-LM were correct (unpublished data). In addition, Johnson et al. 2014 reported an AKH-like neuropeptide in this same species. Also reported are evolutionary relationships between AKHs with Corazonin (CRZ) characterized in American cockroach in 1989 (Veenstra, 1989) and mammalian Gonadotropin-Releasing Hormone (GnRH) (Morgan and Millar, 2004), which was isolated and characterized in invertebrates such as, for example, the common octopus Octopus vulgaris (Iwakoshi et al. 2002). Subsequently, Hansen et al. (2010) described peptide precursors in insects named AKH/Corazonin-related peptide (ACP) but they did not have the proposed NPP organization. That is, one copy of AKH and other for Corazonin within the precursor.
In 2019, together with my student Bachelor of Science Cristian E. Cadena-Caballero we decided to include the precursors of Corazonin, ACP and GnRH in the model. In addition, in the search for APGW/RPCH or AKH NPPs, we collaborated with Ph.D. Laura Rebeca Jimenez-Gutierrez, who was capturing invertebrates in the Gulf of California to obtain transcriptomes and characterize the neuropeptide precursors related to the fishery and reproduction of crustaceans of commercial interest. Among them, the wild crabs Callinectes toxotes and Callinectes arcuatus, of which, we characterized the precursors of ACP and partial CRZ, respectively.
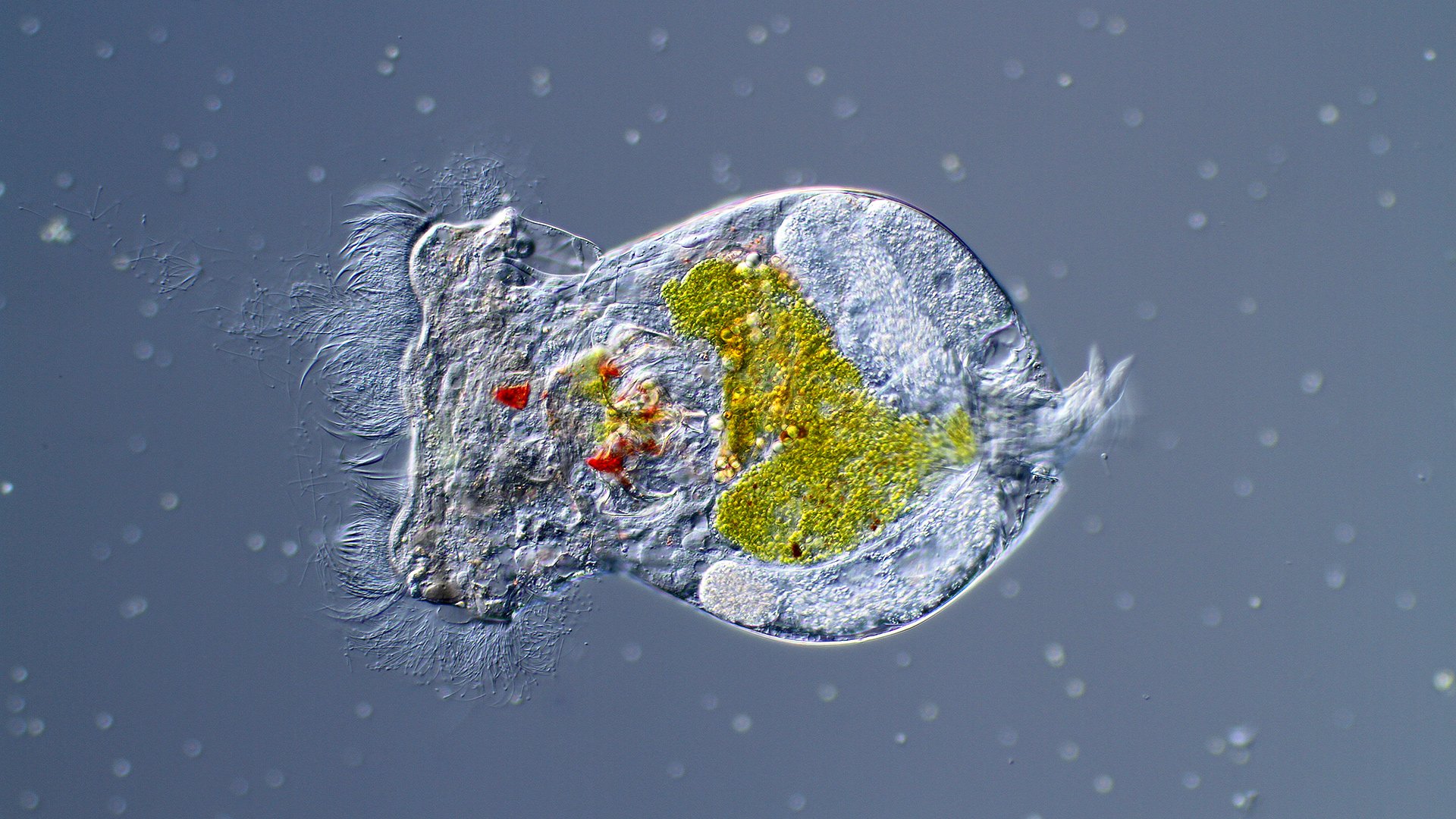
The search for virtual peptides was surprisingly concluded by BLAST in the genome of the rotifer Brachionus plicatilis which reported a hypothetical protein, but in reality, it was the virtual precursor APGW/AKH generated by the loss of codons in the APGWamdie from A. californica and L. stagnalis. Furthermore, this event also occurs in the transcriptomes of other invertebrates that were named with the same name or other APGWs, RPCH, AKH or brain peptide that included the word "like", when in reality they were neuropeptide precursors similar to those described in 2007, by codon fusion of the molluscan APGW precursors (Cadena-Caballero et al., 2023).
We in this publication show to the community the effort and quest of more than 20 years of research to understand neuropeptide evolution by demonstrating for the first time that Neuropeptide Precursor Predictive precursors predicted 22 years before this year, exist in the genome of microscopic animals from the wildlife. That is, precursors with copies of APGW from mollusks and RPCH/AKH from crustaceans or insects. This gives new insight into the evolution of neuropeptide families by DNA-LM and highlights the contributions of all the scientists who initiated the basic science of neuroscience in the 20th century.
Follow the Topic
-
Journal of Molecular Evolution
This journal covers experimental, computational, and theoretical work aimed at deciphering features of molecular evolution and the processes bearing on these features.



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in