The many faces of endothelial cells in healthy and malignant breast tissue – from angiogenesis to immunoregulation and lipid processing
Published in Cancer
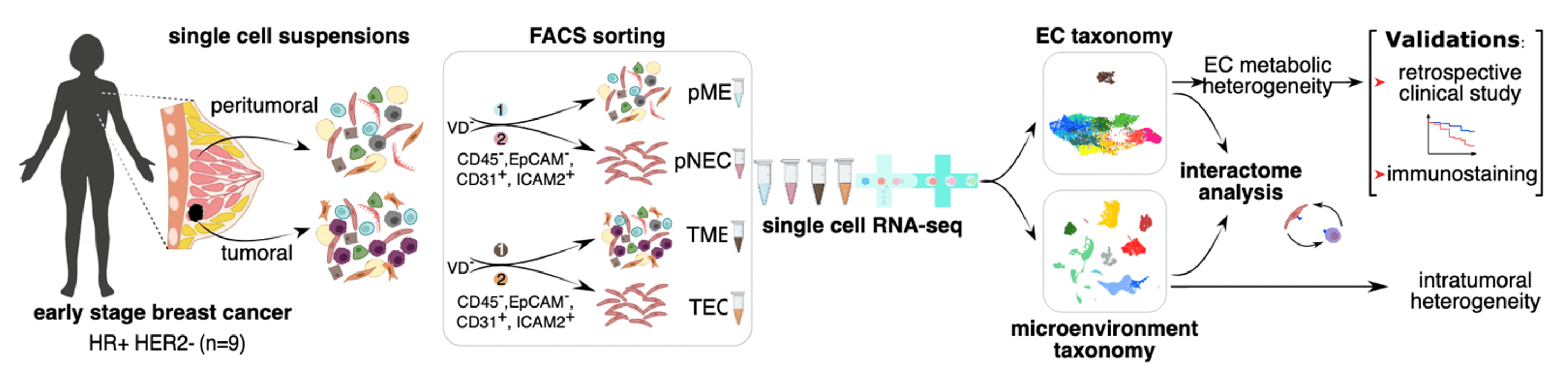
In many aspects, breast cancer (BC) represents a heterogeneous malignancy that globally affects millions of women. This heterogeneity, both within and between patients, has been repeatedly used for prognostic, diagnostic and therapeutic purposes1-3. Different cellular subtypes present within the tumor microenvironment (TME) also display notable heterogeneity, and single-cell RNA-sequencing (scRNA-seq) studies have vastly accelerated our view on stromal cellular subtype diversity (e.g., immune cells, cancer cells and fibroblasts) in different BC patients and subtypes4-10. While the vasculature represents an essential member of the TME, the characterization of vascular heterogeneity in BC, however, has not extended beyond the traditional angiogenic, arterial, capillary, venous and lymphatic endothelial cells (ECs). Yet, obtaining a better understanding of the different EC subtypes that make up the breast (tumor) vasculature, as well as their relation to the TME, represents a pressing but unmet need, as tumor blood vessels are active contributors to tumor growth, invasion and metastasis. Moreover, while current anti-angiogenic therapies (AATs) are indiscriminately used to prune blood vessels in diverse cancer types including BC, these therapies suffer from resistance and insufficient efficacy, and elucidating EC heterogeneity may thus open up opportunities for the development of future novel anti-angiogenic targets.
One of the main reasons why vascular heterogeneity has remained under-characterized in BC thus far, despite the availability of numerous scRNA-seq studies in human BC published to date, is the fact that the abundance of the vascular compartment within individual studies is often relatively low. Moreover, those studies that captured large(r) fractions of ECs generally lack non-malignant control tissue9,10. For both reasons, a detailed and rigorous interrogation of endothelial heterogeneity in human breast tissue, as well as an in-depth comparison of EC subtypes between healthy and malignant tissue, has been lacking thus far from the literature.
To that end, we collected tumor tissue and matched peri-tumoral (non-malignant, as far away from the primary tumor as possible) tissue from 9 BC patients. All patients were treatment-naïve and diagnosed with early, hormone receptor positive/HER2-negative cancers (Figure 1), thus representing one of the most frequent BC types11. We FACS-sorted single cells into two fractions: (1) Viability Dye–/CD45–/EpCAM–/CD31+/ICAM2+ cells, representing the EC-enriched fraction, and (2) Viability Dye– cells without any further selection/enrichment, representing the full TME (Figure 1). Both fractions were subjected to scRNA-seq, and while the former fraction was used to generate a single-cell taxonomy of the BC vasculature, the latter was used to study cell-cell communication between ECs and their neighboring cell types, including immune cells (Figure 1).
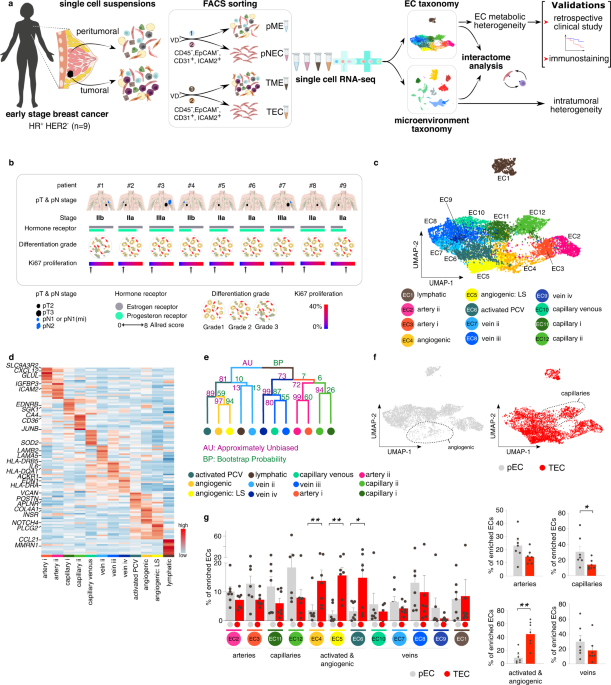
In the EC-enriched arm of our study we obtained 8,433 high-quality ECs that could be categorized into 12 transcriptomically distinct subtypes. We identified all usual suspects, including traditional arteries, capillaries, veins and lymphatic ECs. However, likely given the high resolution of our study, we managed to even go beyond this traditional classification by dividing arterial and capillary ECs into two, and veins into four different subsets with their own set of high-ranking marker genes. We moreover identified multiple subclusters expressing genes known to be enriched in angiogenic blood vessel subtypes (e.g. tip and stalk cells, activated post-capillary venules).
When comparing our BC EC taxonomy to a previously published single-cell study of >21,000 enriched ECs in human lung cancer12, we revealed that most ECs in human breast and lung (cancer) tissue could be divided into similar subtypes. Lymphatic and angiogenic EC subtypes were particularly similar between both tumor types, and especially the latter finding may be interesting in the search for novel, pan-cancer anti-angiogenic candidates. Nevertheless, a fraction of breast (cancer) ECs appeared transcriptionally distinct from human lung (cancer) tissue. We showed that most ECs unique to human breast were of microvascular nature. In fact, they belonged to a specific capillary subtype which will be described in more detail below.
Veins also showed notable differences when comparing their transcriptomes in breast and lung (cancer) ECs. Notably, several genes involved in tumor immunity and/or antigen presentation were enriched in the different BC vein EC subclusters, suggesting a potential role for veins in shaping the BC microenvironment. Further receptor-ligand interaction analysis (including the non-EC-enriched arm of our study) revealed numerous interactions between ECs and immune cells that further corroborated our hypothesis that (venous) ECs might play immunomodulatory roles in BC. For instance, we predicted venous ECs to interact closely with cytotoxic natural killer (NK) cells via communication through the CLEC2B-KLRF1 axis. We confirmed this interaction in situ, and furthermore showed that knock-down of CLEC2B in human umbilical vein ECs was able to reduce the percentage of freshly isolated, activated human NK cells in co-culture experiments, thus extending our in silico predicted interaction to a functional role for ECs in NK cell activation.
Lastly, we revealed a distinct capillary EC subtype enriched in peri-tumoral breast tissues, not identified in lung (cancer), and expressing genes involved in various flavors of lipid metabolism (e.g. lipid uptake, lipid droplet turnover, transcription factor-mediated regulation of fatty acid metabolism (e.g. PPARG)). Hence, we referred to these capillaries as Lipid Processing ECs (LIPECs), and confirmed their increased presence in peritumoral BC tissue using immunohistochemistry and/or in situ hybridization (RNA-scope). Retrospectively, we furthermore showed that PPARG signaling activation, indirectly through intake of the biguanide metformin (one of the agents used as standard of care treatment of diabetes mellitus13,14) was associated with reduced mortality and metastases in BC patients. Moreover, the abundance of LIPECs was significantly increased in tumors from metformin-treated BC patients compared to those from patients who did not receive this treatment. While the contribution of LIPECs to BC and the precise mechanisms of how metformin or PPARG regulates lipid metabolism in LIPECs requires further study, the prognostic relevance of LIPEC abundance as well as the clinical benefit of stimulating their presence or abundance present intriguing concepts to be explored in the near future.

Contributors: Laura de Rooij and Peter Carmeliet
References:
1 Turashvili, G. & Brogi, E. Tumor Heterogeneity in Breast Cancer. Front Med (Lausanne) 4, 227-227, doi:10.3389/fmed.2017.00227 (2017).
2 Koren, S. & Bentires-Alj, M. Breast Tumor Heterogeneity: Source of Fitness, Hurdle for Therapy. Molecular Cell 60, 537-546, doi:10.1016/j.molcel.2015.10.031 (2015).
3 Harbeck, N. et al. Breast cancer. Nature Reviews Disease Primers 5, 66, doi:10.1038/s41572-019-0111-2 (2019).
4 Azizi, E. et al. Single-Cell Map of Diverse Immune Phenotypes in the Breast Tumor Microenvironment. Cell 174, 1293-1308 e1236, doi:10.1016/j.cell.2018.05.060 (2018).
5 Bartoschek, M. et al. Spatially and functionally distinct subclasses of breast cancer-associated fibroblasts revealed by single cell RNA sequencing. Nat Commun 9, 5150, doi:10.1038/s41467-018-07582-3 (2018).
6 Chen, W. et al. Single-cell landscape in mammary epithelium reveals bipotent-like cells associated with breast cancer risk and outcome. Commun Biol 2, 306, doi:10.1038/s42003-019-0554-8 (2019).
7 Chung, W. et al. Single-cell RNA-seq enables comprehensive tumour and immune cell profiling in primary breast cancer. Nature communications 8, 15081-15081, doi:10.1038/ncomms15081 (2017).
8 Savas, P. et al. Single-cell profiling of breast cancer T cells reveals a tissue-resident memory subset associated with improved prognosis. Nature Medicine 24, 986-993, doi:10.1038/s41591-018-0078-7 (2018).
9 Wu, S. Z. et al. A single-cell and spatially resolved atlas of human breast cancers. Nature Genetics 53, 1334-1347, doi:10.1038/s41588-021-00911-1 (2021).
10 Bassez, A. et al. A single-cell map of intratumoral changes during anti-PD1 treatment of patients with breast cancer. Nature Medicine 27, 820-832, doi:10.1038/s41591-021-01323-8 (2021).
11 Meisel, J. et al. Evaluation of Prognosis in Hormone Receptor-Positive/HER2-Negative and Lymph Node-Negative Breast Cancer With Low Oncotype DX Recurrence Score. Clin Breast Cancer 18, 347-352, doi:10.1016/j.clbc.2017.12.006 (2018).
12 Goveia, J. et al. An Integrated Gene Expression Landscape Profiling Approach to Identify Lung Tumor Endothelial Cell Heterogeneity and Angiogenic Candidates. Cancer Cell 37, 21-36.e13, doi:https://doi.org/10.1016/j.ccell.2019.12.001 (2020).
13 Gross, B. & Staels, B. PPAR agonists: multimodal drugs for the treatment of type-2 diabetes. Best Practice & Research Clinical Endocrinology & Metabolism 21, 687-710, doi:https://doi.org/10.1016/j.beem.2007.09.004 (2007).
14 Hong, F., Xu, P. & Zhai, Y. The Opportunities and Challenges of Peroxisome Proliferator-Activated Receptors Ligands in Clinical Drug Discovery and Development. Int J Mol Sci 19, 2189, doi:10.3390/ijms19082189 (2018).
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026

![When PSMA-targeted therapy is not enough: high-risk localized prostate cancer after repeated [177Lu]Lu-PSMA radioligand therapy](/cdn-cgi/image/metadata=copyright,fit=scale-down,format=auto,quality=95,width=256,height=256/https://public-storage.zapnito.com/Ku6h7Yyp4Q0LXqRRMICCHR2v4LcOsmxMrmDPtOYuI1c)



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in