The story of OptoMedicine: REDLIP system for metabolic homeostasis control
Published in Bioengineering & Biotechnology

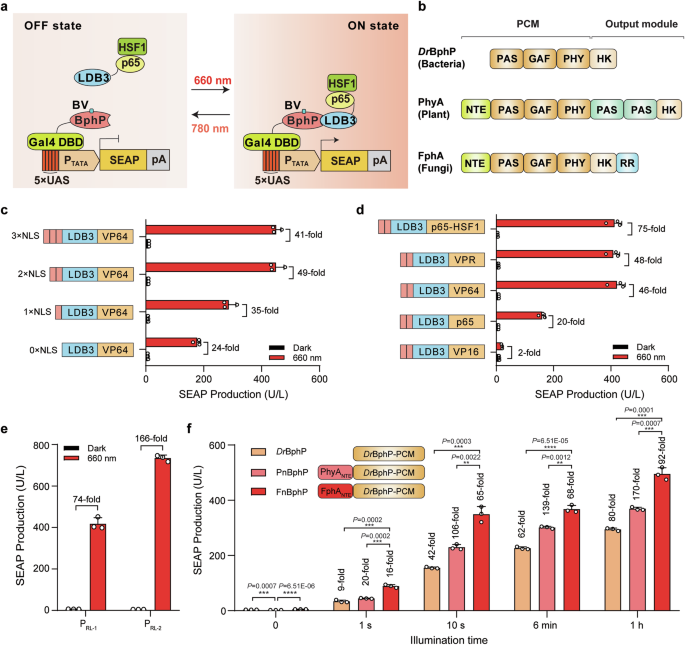
Light has some features including high spatiotemporal resolution, excellent adjustability and reversibility, and optogenetics has been widely used for the light-mediated manipulation of biological processes in living organisms, which has paved the way for precision interventions in gene- and cell-based therapies1, 2. Our laboratory has focused on developing novel optogenetic tools for biomedical applications. In 2011, Dr. Haifeng Ye designed a blue light-responsive optogenetic switch to induce shGLP1 production for regulating blood glucose homeostasis in a diabetic mouse model3. However, the genetic circuit's nonorthogonality and the limited tissue penetration of blue light constrained its broader application.
Red/far-red light is preferable to other wavelengths4, 5, due to its advantages including deep tissue penetration, low light scattering, and low photo-cytotoxicity2. In 2017, our group developed a far-red light-controlled transgene expression system based on BphS5 to regulate gene expression. This system was further integrated with CRISPR/Cas9, CRISPR/dCas9, and Cre recombinase, enabling precise control of epigenome engineering6, gene editing7, and DNA recombination8 both in vitro and in vivo. However, this system had some limitations, including the need for prolonged illumination to achieve full activation and the complexity of its relatively large genetic modules, which posed challenges for packaging into adeno-associated virus (AAV) vectors for gene therapy.
In 2021, our group developed a red/far-red light-mediated and miniaturized Δphytochrome A (ΔPhyA)-based photoswitch system (REDMAP)4, which is characterized by high transcriptional activation efficiency (> 150-fold) and rapid activation/deactivation (~1 s) kinetics. This system was further integrated with the endogenous Ras/Erk mitogen and CRISPR/dCas9, enabling precise control of protein kinase (MAPK) activating and epigenome engineering. However, its applicability, especially for long-term controllable gene therapy, is still limited by the need for phycocyanobilin (PCB), which must be provided through exogenous administration or engineering cells to express PCB biosynthesis enzymes.
To overcome these shortcomings, we developed a red/far-red light-inducible photoswitch (REDLIP) system based on the chimeric photosensory protein PnBphP or FnBphP and their interaction partner LDB39 (Fig. 1). PnBphP or FnBphP was engineered by fusing N-terminal extension (NTE) of phytochrome A (PhyA) or fungal phytochrome FphA derived from Aspergillus nidulans to N-terminal DrBphP-PCM10, 11, where PhyA’s NTE was reported to influence photoresponse properties by stabilizing the active Pfr state12. These genetically encoded REDLIP systems require no external biliverdin, exhibit fast ON/OFF kinetics (10 seconds red light illumination and 1-minute far-red light illumination), strong activation of target gene expression (> 100-fold), and a dose-dependent response to red/far-red light.

Fig. 1 Graphical abstract for this study
After successfully demonstrating that the REDLIP system could induce highly efficient exogenous gene transcription under red light illumination, we sought to explore its potential for controlling endogenous gene activation. To this end, we developed the REDLIP-mediated CRISPR-dCas9 system (REDLIPcas), which enabled a remarkable induction of multiple endogenous genes—over 1100-fold—under just 10 seconds of red light exposure.
One of the key advantages of the REDLIP system is its compact size, which allows it to be packaged into AAV vectors, making it suitable for therapeutic applications. To demonstrate its potential in treating metabolic diseases, we used an AAV-delivered Fn-REDLIP system to optogenetically control insulin expression, with 30-minute twice-daily light exposure effectively lowering blood glucose levels in a type 1 diabetes mouse model (Fig. 2a). Additionally, we showed that the system regulated thymic stromal lymphopoietin (TSLP) expression to normalize body weight in obesity mouse models after 30-minute light exposure every three days for four weeks (Fig. 2b).

Fig. 2 AAV-delivered REDLIP-mediated gene therapy in type 1 diabetes and obesity mouse models.
In designing practical applications for the REDLIP system, we also focused on minimizing unwanted activation from ambient light. To address this challenge, we conducted in vivo experiments where AAV-delivered REDLIP mediated TSLP transgene expression in the muscles of high-fat diet (HFD) mice under the control of an LED patch, operated by a smartphone (Fig. 3). This setup demonstrated that the LED patch could precisely control gene expression on demand without unwanted ambient light activation, offering a promising strategy for treating diseases.
Fig. 3 AAV-delivered REDLIP-mediated expression of TSLP in the muscles of HFD mice controlled by an LED patch operated via smartphone.
In conclusion, the REDLIP system represents a powerful optogenetic tool capable of efficiently inducing gene expression in mammals without the need for external chromophores. The system operates on a rapid timescale of seconds, allows for deep tissue penetration, and is minimally invasive. This light-controlled gene therapy strategy shows potential for the precise treatment of diseases requiring periodic regulation of hormone-based drugs, such as diabetes, thyroid disorders, and menstrual cycle-related hormonal imbalances. With its potential for therapeutic applications, we believe the AAV-delivered REDLIP system will accelerate the clinical adoption of optogenetic therapies, opening new possibilities for the treatment of metabolic diseases and beyond.
Our paper (also ref. 9 below): Qiao, L. et al. A sensitive red/far-red photoswitch for controllable gene therapy in mouse models of metabolic diseases. Nat. Commun. (2024). https://doi.org/10.1038/s41467-024-54781-2
References
- Bansal, A. et al. Towards translational optogenetics. Nat. Biomed. Eng. 7, 349-369 (2023).
- Tan, P. et al. Optophysiology: Illuminating cell physiology with optogenetics. Physiol. Rev. 102, 1263-1325 (2022).
- Ye, H. et al. A synthetic optogenetic transcription device enhances blood-glucose homeostasis in mice. Science. 332, 1565-1568 (2011).
- Zhou, Y. et al. A small and highly sensitive red/far-red optogenetic switch for applications in mammals. Nat. Biotechnol. 40, 262-272 (2022).
- Leopold, A.V. et al. A general approach for engineering RTKs optically controlled with far-red light. Nat. Methods. 19, 871-880 (2022).
- Shao, J. et al. Synthetic far-red light-mediated CRISPR-dCas9 device for inducing functional neuronal differentiation. Proc. Natl Acad Sci. U S A. 115, E6722-e6730 (2018).
- Yu, Y. et al. Engineering a far-red light–activated split-Cas9 system for remote-controlled genome editing of internal organs and tumors. Sci. Adv. 6, eabb1777 (2020).
- Wu, J. et al. A non-invasive far-red light-induced split-Cre recombinase system for controllable genome engineering in mice. Nat. Commun. 11, 3708 (2020).
- Qiao, L. et al. A sensitive red/far-red photoswitch for controllable gene therapy in mouse models of metabolic diseases. Nat. Commun. 15, 10310 (2024).
- Nagano, S. et al. Structural insights into photoactivation and signalling in plant phytochromes. Nat. Plants. 6, 581-588 (2020).
- Brandt, S. et al. The fungal phytochrome FphA from Aspergillus nidulans. J. Biol. Chem. 283, 34605-34614 (2008).
- Gourinchas, G. et al. Influence of the N-terminal segment and the PHY-tongue element on light-regulation in bacteriophytochromes. J. Biol. Chem. 294, 4498-4510 (2019).
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Biosensing
Publishing Model: Hybrid
Deadline: Sep 30, 2026





Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in