When do polyploidy and self-compatibility go hand in hand?
Published in Ecology & Evolution


Polyploidy or whole genome duplication, often accompanied by hybridization, is a common speciation mode in flowering plants. But newly formed polyploids immediately face a problem – a lack of compatible mates of the same ploidy. If the new polyploids are obligately outcrossing due to genetic self-incompatibility, as many plants are, this problem is exacerbated. For these reasons, researchers have long hypothesized that self-compatibility, the ability to produce seeds by self-fertilization, should be more common in polyploids, as it would increase their chances of successful establishment. However, systematic analyses have found mixed evidence for such an association, with the exception of some plant families.
In our recent study in Heredity, PhD student Jörg Bachmann with colleagues at Stockholm University and SLU set out to test whether polyploidy could sometimes lead to instant self-compatibility in the mustard family. We suspected that such an association could be an effect of the particular molecular details of the genetic system for self-incompatibility. In the Brassicaceae, dominant alleles at the self-incompatibility locus (S-locus) can suppress the expression of recessive alleles in pollen. Presence of a dominant non-functional S-allele could thus lead to instant self-compatibility, especially in allopolyploids that form by hybridization and genome duplication (Novikova et al. 2017). In this way, polyploidy and self-compatibility could sometimes go hand in hand.
The widespread weed Shepherd’s Purse (Capsella bursa-pastoris) is an interesting species in which to investigate this scenario. This very common weedy species is an allotetraploid that formed by hybridization and polyploidization about 200-300 kya, and the closest extant relatives of its parental species come from the C. orientalis and C. grandiflora lineages, respectively (Douglas et al. 2015).

Because C. orientalis is self-compatible, we hypothesized that if its S-allele was dominant, self-compatibility could have been instant upon the formation of C. bursa-pastoris. Simply put, a non-functional but dominant S-allele from C. orientalis could repress the expression of a functional allele inherited from the other progenitor, resulting in instant self-compatibility. In the mustard family such dominance relationships among S-alleles are common and often mediated by small RNAs.
To test this hypothesis we sequenced the entire S-locus in both subgenomes of C. bursa-pastoris and searched for small RNA-based candidate dominance modifiers in these sequences. We found a shared loss-of-function mutation between C. bursa-pastoris and C. orientalis at SCR, a key gene for self-incompatibility, indicating that the S-allele from C. orientalis was likely ancestrally non-functional in C. bursa-pastoris. The S-allele inherited from C. orientalis further expressed a candidate sRNA-based dominance modifier that shows sequence conservation over 20 million years. Together, these findings suggest that a non-functional S-allele inherited from C. orientalis dominantly suppressed the S-allele inherited from the other progenitor. This suggests that self-compatibility might have been immediate upon formation of the tetraploid Shepherd’s Purse, helping establishment of the new species.
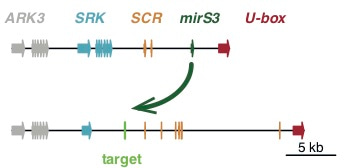
In our paper we also show empirically that such instant self-compatibility is possible in Capsella. This was in fact one piece of evidence that we did not originally set out to collect. Rather, as part of a different study, Jörg unexpectedly obtained tetraploid self-compatible offspring from F1 hybrids of C. orientalis and C. grandiflora. We then screened the F1 hybrids and found evidence for somatic doubling involving flowering branches. The fact that we found tetraploid self-compatible offspring demonstrates that instant self-compatibility upon allopolyploidization is possible, but the outcome would of course depend on the specific combination of S-alleles and their respective dominance relationships.
Interestingly, there are other cases described in the literature of allotetraploids that have formed this way – one classic example being Primula kewensis which formed after a wide cross between Primula verticillata and Primula floribunda (Newton and Pellew 1929). Whether C. bursa-pastoris originally formed through somatic doubling after wide hybridization is still unclear, but based on our findings we suggest that this pathway to polyploidy should at least be considered.



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in
Great study - finally we can understand (at least in this case) how polyploidy and the ability to self can evolved at the exact same moment.