Brain Networks of Social and Asocial Learning
Published in Neuroscience

The social intelligence hypothesis suggests that living in social groups leads to the development of enhanced cognitive abilities to cope with the complexity and unpredictability of social interactions (1,2). This drives the evolution of more complex cognitive skills and favors the growth of larger executive brains (3,4). There are two proposed scenarios for how cognitive abilities can be encoded in the brain: general-purpose or the special-purpose. The general-purpose suggests that cognitive mechanisms for information processing, such as input, encoding, storage, and retrieval, are shared across different functional domains. This means that enhanced cognitive abilities evolved in a specific domain, such as social interactions, can also be advantageous in other domains like foraging and predator avoidance (5–9). The special-purpose proposes that each functional domain relies on specialized cognitive modules with independent information processing mechanisms (5–9). Another perspective suggests that social learning and asocial learning operate through the same general learning mechanisms, with specialized adaptations primarily present in the input systems (6–9).
Research on the genetic and neural mechanisms of social and asocial learning can clarify the presence, or not, of shared processes. Several studies used specific brain regions that are chosen a priori as candidates for social learning tasks (10–15), and studies using an unbiased brain network approach are lacking in this field. Additionally, localized neuronal activation analyses do not reveal the patterns of co-activation across brain networks, which could uncover specialized or conserved modules for different learning types.
In this study, a classic conditioning paradigm in zebrafish was employed to investigate the neural basis of social and asocial learning. Zebrafish were exposed to either a social stimulus (image of a zebrafish) or an asocial stimulus (image of a circle) paired with a food reward. The choice of static images of fish as a social stimulus was based on the known attraction elicited by biological motion and body shape/coloration patterns. The study utilized the expression of the immediate early gene c-fos as a marker of neuronal activity during retrieval. A new method was developed to analyze brain network functional connectivity using co-activation matrices and network community analysis was employed to identify coherent sets of brain regions consistently recruited during social and asocial learning tasks.
Zebrafish demonstrated equal learning abilities when exposed to either social or asocial-conditioned stimuli (CS) paired with an unconditioned stimulus (US). There were no significant differences between the social and asocial learning treatments in learning acquisition during the training phase or retrieval during the probe test. Importantly, zebrafish were able to distinguish between the social and asocial cues used in the study, and they showed a preference for the social cue.
The levels of brain activation, measured by c-fos expression, revealed distinct patterns for the retrieval of social and asocial learning. Social learning was associated with increased activity in the olfactory bulbs (OB), the ventral zone of the ventral telencephalic area (Vv), the ventral habenular nuclei (Hav), and the ventromedial thalamic nuclei (VM). In contrast, asocial learning was associated with decreased activity in the dorsal habenular nuclei (Had) and in the anterior tubercular nucleus (ATN).
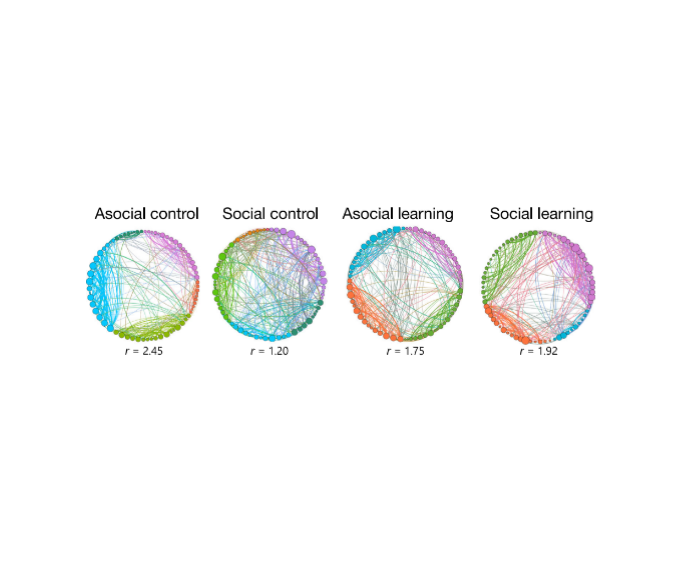
The regional similarity data reveals a lack of correlation between social and asocial learning, and a large amplitude of the change in the neighborhood structure for different regions in the two types of learning. Community detection analysis revealed a higher integration across communities in the social learning treatment compared to the asocial one. Additionally, this study identifies shared submodules within the brain networks. Control groups share the visual response to a stimulus but include no learning on the part of the individuals: generalized attention (AM) and the visual response mechanisms (VRM). Social learners compared with social controls, the commonality lies in the presence of a social visual stimulus, composed of VRM regions and a few additional ones, that we now denote as social integration module (SIM). In asocial learners compared with asocial controls, the submodule includes about 50% of the VRM regions plus a new set of regions with no overlap with the SIM. Arguably, these additional regions should be responsible for the learned response and association with the food stimulus, and we denote them as the learning module (LM). The region VL l, which appears in all the submodules, might suggest that it has a generic role in information integration across different areas. In our study, the social module is exclusively composed of brain regions present in the right hemisphere. Similarly, the learning module is also lateralized, but in this case, it is mostly composed of left-hemisphere structures. These results suggest the occurrence of laterality in zebrafish brain function, which may increase efficiency in information processing by avoiding duplication of functions in the two hemispheres.
Together, our results provide the first experimental evidence for the occurrence of a general-purpose learning module that is apparently modulated by different patterns of localized activity in social and asocial learning.
https://doi.org/10.1038/s42003-023-04999-5
References
- Byrne, R.W. & Whiten, A. Machiavellian intelligence: social expertise and the evolution of intellect in monkeys, apes, and humans (Oxford, 1988).
- Byrne, R. W. & Bates, L. A. Sociality, evolution and cognition. Curr. Biol. 17, R714–R723 (2007).
- Dunbar, R. I. M. The Social brain hypothesis. Evol. Antropol. 6, 178–190 (1998).
- Dunbar, R. I. M. & Shultz, S. Evolution in the social brain. Science 317, 1344–1347 (2007).
- Heyes, C. M. Social learning in animals: categories and mechanisms. Biol. Rev. Camb. Philos. Soc. 69, 207–231 (1994).
- Heyes, C. What’s social about social learning? J. Comp. Psychol. 126, 193–202 (2011).
- Heyes, C. & Pearce, J. M. Not-so-social learning strategies. Proc. Biol. Sci. 282, 20141709 (2015).
- Leadbeater, E. What evolves in the evolution of social learning? J. Zool. 295, 4–11 (2014).
- Varela, S. A. M., Teles, M. C. & Oliveira, R. F. The correlated evolution of social competence and social cognition. Funct. Ecol. 34, 332–343 (2020).
- Olsson, A. & Phelps, E. A. Social learning of fear. Nat. Neurosci. 10, 1095–1102 (2007).
- Jeon, D. et al. Observational fear learning involves affective pain system and Cav1.2 Ca2+ channels in ACC. Nat. Neurosci. 13, 482–488 (2010).
- Allsop, S. A. et al. Corticoamygdala transfer of socially derived information gates observational learning. Cell 173, 1329–1342.e18 (2018).
- Burgos-Robles, A., Gothard, K. M., Monfils, M. H., Morozov, A. & Vicentic, A. Conserved features of anterior cingulate networks support observational learning across species. Neurosci. Biobehav. Rev. 107, 215–228 (2019).
- Carrillo, M. et al. Emotional mirror neurons in the rat’s anterior cingulate cortex. Curr. Biol. 29, 1301–1312.e6 (2019).
- Munger, S. D. et al. An olfactory subsystem that detects carbon disulfide and mediates food-related social learning. Curr. Biol. 20, 1438–1444 (2010).
Follow the Topic
-
Communications Biology
An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.

Related Collections
With Collections, you can get published faster and increase your visibility.
Artificial Intelligence Methodology in Structural Biology
Publishing Model: Hybrid
Deadline: Nov 30, 2026
Healthy Aging
Publishing Model: Open Access
Deadline: Dec 31, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in