Deep eukaryotic roots for the underground fungal network.
Published in Ecology & Evolution

Fungal filaments, hyphae, are one of the defining traits of fungi that, among others enable them to forage for nutrients, find mating partners and even maintain a surprisingly low mutation rate, as shown recently. The intricate network of hyphae form a multicellular thallus, called mycelium. Thin hyphal filaments can invade food sources efficiently and are considered a main virulence factor for pathogenic fungi, especially for dimorphic fungi that can switch between unicellular yeast and multicellular hyphal forms.
Hyphae-like structures occur only in a few groups, including oomycetes, actinobacteria and, obviously, fungi. These filaments grow apically by tip extension, making them the most polarized cells in nature. Hyphae set fungi apart from other groups, but what sets hyphae apart from other forms of multicellularity?
Researchers have been pondering this question for a long time, but attempts to unequivocally categorize filamentous fungi in the realm of multicellular forms have proved difficult. Most multicellular organisms seem to quite seamlessly fit into two main modes. Clonal multicellularity refers to colonies formed by descendants of a founder cell (e.g. by daughter cells failing to separate from mother cell). Aggregative multicellularity arises when cells of varying relatedness aggregate and form one higher level entity, such as seen in Dictyostelium slime molds. In a typical filamentous fungus, e.g. one that belongs to the Dikarya, cells are formed by closing hyphal sections off by septa, with regulated partitioning of nuclei between the newly separated and the apical cell. Although at the level of nuclei this mechanism resembles clonal multicellularity, it isn't exactly that. Observations on the organization of multicellular growth in fungi suggested that hyphae might represent a unique way to evolve multicellularity.
If the mechanism of multicellular growth in fungi differs so much from that of other multicellular organisms, how do mechanisms of its emergence and the principles that govern multicellularity in fungi compare? Are the principles we learned by examining canonical multicellular model systems, such as Volvox, choanoflagellates or Dictyostelium, apply to fungi?
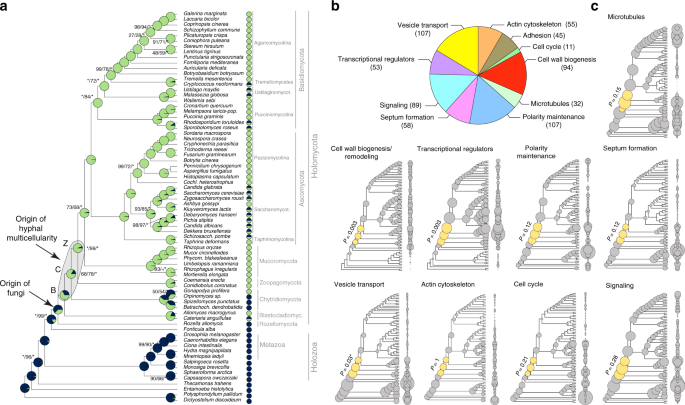
In our recent paper, we took a comparative genomic approach to reveal the evolutionary origins of hyphal growth. Fortunately, mycologists have been studying the genetics and cell biology of hyphal growth for ages, so plenty of genes were already known that participated in the process. Enikő Kiss, a PhD student in the lab at the time, painstakingly searched the mycological literature for genes associated with hyphal growth. After going through hundreds of papers we finally concluded that likely much of the knowledge on hyphal growth was integrated into her database. After going through the detected genes, we restricted the analysis to ones that are directly connected to hyphal growth i.e. necessary and sufficient for a pathway to function - some 650 genes.
We identified the gene families that contained these genes and analyzed their evolution using a phylogenetic approach. These analyses revealed some surprises, including the low diversity of G-protein coupled receptors, kinases and adhesion-related genes in the most recent common ancestor of all filamentous fungi. Multicellular lineages like animals and plants show expansions in these gene families, whereas in fungi it appears the awaited expansion is missing. Where are then the missing receptors in fungi? Maybe they are not G-protein coupled receptors or we haven't discovered them yet?
What appears to be shared by fungi and other multicellular lineages is a deep conservation of multicellularity-related genes. Groundbreaking works of Nicole King's and Iñaki Ruiz-Trillo's lab, among others, early on showed that several key metazoan signaling and adhesion pathways are conserved in choanoflagellates. In fungi, many of the cellular systems involved in hyphal growth - vesicle trafficking, cytoskeleton and polarity maintenance, among others - predate the origin of the first filamentous fungi. This indicates that, rather than the evolution of new genes, the co-option and exaptation of ancient genes dominates the evolutionary landscape of hyphal origins. A striking example of provided by phagocytosis-related genes. Fungal cells - with a rigid cell wall - are unable to phagocytose, yet we found many phagocytosis-related genes are conserved in fungi (this was also noted earlier by Torruella et al). In fungi, these 'surviving' phagocytotic genes are involved in recycling excess membrane material that forms as a result of continuous apical secretion vesicles containing cell wall materials. This suggests an exaptation event: their ancestral function was lifted by the emergence of a cell wall (and transition to osmotrophic lifestyle) but were recruited for hyphal growth during evolution.
This work adds to the quickly piling evidence that much of the evolutionary innovation out there, small or major, is based on the rewiring of existing gene networks, rather than dozens or hundreds of new genes.
Image credit: http://www.sclerotia.org/lifecycle/hyphal-growth
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in