in vivo evidence for soluble ACE2 as an entry receptor of SARS-CoV-2
Published in Healthcare & Nursing
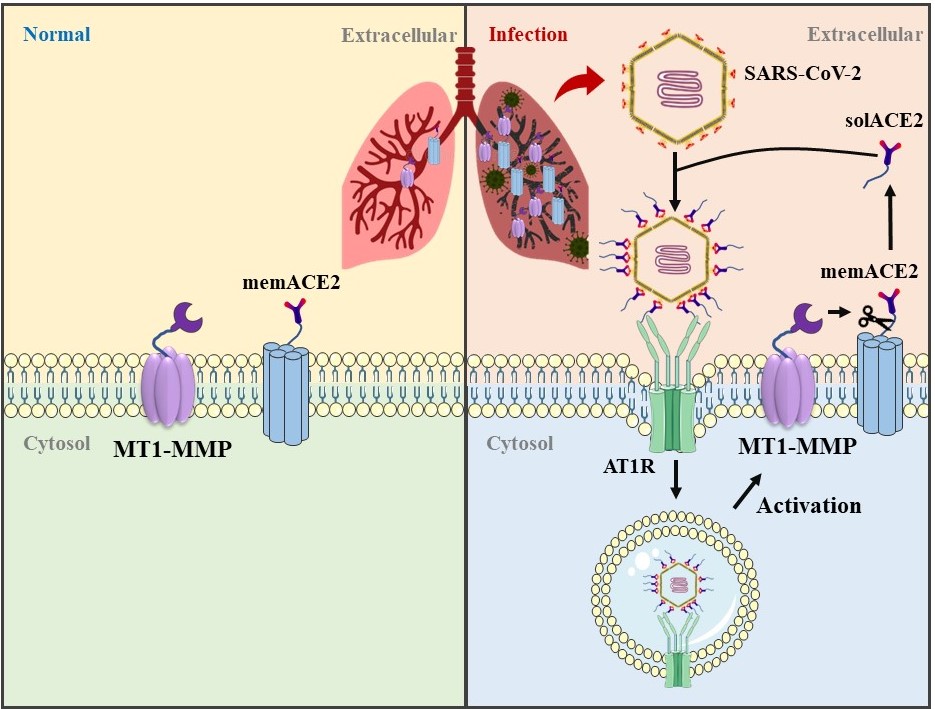

A novel severe acute respiratory syndrome (SARS)-like coronavirus (SARS-CoV-2) is causing the global coronavirus disease 2019 (COVID-19) pandemic with high mortality rate, especially for the elderly population. Understanding the cell entry mechanism of SARS-CoV-2 is of high priority in order to curb its spread. Angiotensin converting enzyme 2 (ACE2), a well-known cell entry receptor of SARS-CoV-2, exists in two major forms: the full-length membrane form (memACE2) and a shed soluble form (solACE2). While the role of memACE2 in mediating SARS-CoV-2 cell entry has been well-documented, the function of solACE2 in SARS-CoV-2 infection remains debating. Under the physiological situation, solACE2 released by cell surface proteases has been found to serve as the cellular doorway to SARS-CoV-2, facilitating receptor-mediated endocytosis of virus1. However, there are conflicting results in the promoting effects of solACE2 on virus entry2,3. While evidence increasingly links the importance of soluble ACE2 with the infectivity of SARS-CoV-24-6, the underlying mechanism which regulates the release of ACE2 from the cell surface has remained elusive. Although the function of solACE2 in SARS-CoV-2 infection has been demonstrated in vitro1, it is not known how the physiological regulation of ACE2 shedding contributes to the etiology of COVID-19 in vivo.
ACE2 is expressed in multiple tissues in addition to the respiratory system and also corelates with their potential susceptibilities to SARS-CoV-2 infection. The expression of ACE2 in nasal epithelium significantly increases with age, explaining why the ageing population is more susceptible to COVID-19 7. This caught my attention as the ongoing research in my lab identified MT1-MMP/MMP14, a cell surface metalloproteinase important for extracellular matrix remodeling and pericellular proteolysis, as a key host factor for multiple age-associated metabolic diseases8,9. Nonetheless, a previous study has reported the contribution of MT1-MMP to influenza-related tissue damages and mortality in rodents10, highlighting its pathological role in viral infections.
Since my PhD study, my laboratory has been working on identifying the diverse roles of MT1-MMP in different physiological and pathological conditions, such as osteogenesis, lymphangiogenesis, immune system development and obesity9,11-13. Given that MT1-MMP has broad spectrum of substrates, we sought to expand our research on the SARS-CoV-2 receptors. In our study, we found that SARS-CoV-2 infection led to increased ACE2 shedding, mediated by a metalloproteinase MT1-MMP. We further found evidence of MT1-MMP activation in human COVID-19 lung pathological samples. Single cell transcriptome analyses revealed that MT1-MMP was highly colocalized with ACE2 in human lung epithelium and MT1-MMP+ACE2+ epithelial cells are highly susceptible to SARS-CoV-2 infection. Mechanistically, MT1-MMP directly cleaved memACE2 at M706-S to release solACE218-706 that bound to the SARS-CoV-2 spike proteins (S), thus facilitating cell entry of SARS-CoV-2 (Fig. 1). Ectopic expression of MT1-MMP promoted SARS-2-S-driven entry by releasing solACE218-706. Ablation of MT1-MMP cleavage site effectively abolished MT1-MMP-mediated release of solACE2 and MT1-MMP-induced increase in viral entry, which confirms that MT1-MMP promotes SARS-CoV-2 entry by releasing solACE218-706. To further explore the therapeutic potential of targeting MT1-MMP in the treatment of COVID-19, we demonstrated that inhibition of MT1-MMP activity by either genetic knockdown or pharmacological antagonism effectively suppressed solACE218-706 release and attenuated the entry of SARS-CoV-2 and its variants of concern in multiple human primary cells and organoids, including human bronchial epithelial cells, primary human bronchial epithelial 3D cultures with an air-liquid interface (ALI), human cardiac organoids formed by iPSC-derived cardiomyocytes and human liver organoids. To further demonstrate the physiological role of solACE218-706 for SARS-CoV-2 infectivity, we generated a mouse model expressing human solACE218-706 by adenoviral transduction. In comparison with wild-type C57BL/6 mice that were insusceptible to SARS-CoV-2 infection even with high infectious dose, mice expressing human solACE218-706 sustained high viral loads in lung upon intranasal infection of SARS-CoV-2, which confirms the essentiality of solACE2 for SARS-CoV-2 infectivity. More importantly, we demonstrated that the level of sACE2 was significantly elevated in mice and human subjects with ageing, a major risk factor of SARS-CoV-2 infection, and there was a significantly positive correlation between solACE2 and plasma MT1-MMP. Notably, in aged mice infected with mouse-adapted SARS-CoV-2, we found that inhibition of MT1-MMP activities with specific blocking antibodies confers a high level of therapeutic effects.

Figure 1. Graphical illustration shows the MT1-MMP-directed release of solACE2 that mediates the entry of SARS-CoV-2.
In our study, we have delineated the detailed mechanism regulating the release of solACE2 by identifying the major host protease responsible for ACE2 shedding. By comparing with other well-known proteases (e.g. MMP2, MMP9, MMP13, ADAM9 and ADAM17) involved in ectodomain-shedding of cell surface proteins, only MT1-MMP was found to efficiently cleave memACE2 and release functionally active solACE218-706. This ACE2 fragment is indeed the dominant solACE detected in both human and mouse tissues, and can be largely depleted by inhibition of MT1-MMP activities. Given the facts that adenoviral transduction of human solACE218-706 enables SARS-CoV-2 infection in wild-type C57BL/6 mice and blockage of ACE2 shedding by MT1-MMP effectively attenuates SARS-CoV-2 entry in multiple types of human organoids ex vivo and rodents in vivo, we are the first to provide the in vivo evidence showing the contribution of ACE2 shedding to the etiology of COVID-19. Despite the respiratory route being dominant in the infection of SARS-CoV-2, only a small population of the lung cells expresses detectable levels of ACE2. However, SARS-CoV-2 has organotropism beyond the respiratory tract, evidenced by the facts that SARS-CoV-2 infection leads to multiorgan injury and high levels of ACE2 can be detected in kidney, heart muscle and small intestine. Our new data provides a proof of concept that SARS-CoV-2 adopts solACE2-mediated machinery to infect cells with the relatively low expression of ACE2 or without ACE2 expression, facilitating multiorgan infections. In summary, our findings show that MT1-MMP-mediated proteolytic processing of the solACE2 is critical for SARS-CoV-2 infection in vivo and suggest that MT1-MMP is a potential therapeutic target for treatment of COVID-19 and emerging SARS-CoV-2 variants of concern.
Overall, our new findings on SARS-CoV-2 entry mechanism provides new insights into the regulation of viral infection, which will improve our understanding of SARS-CoV-2 organotropism and may guide the development of targeted therapies for COVID-19.
Read the full story in Nature Communications.
References
1 Yeung, M. L. et al. Soluble ACE2-mediated cell entry of SARS-CoV-2 via interaction with proteins related to the renin-angiotensin system. Cell 184, 2212-2228 e2212, doi:10.1016/j.cell.2021.02.053 (2021).
2 Batlle, D. et al. Evidence in favor of the essentiality of human cell membrane-bound ACE2 and against soluble ACE2 for SARS-CoV-2 infectivity. Cell 185, 1837-1839, doi:10.1016/j.cell.2022.05.004 (2022).
3 Yeung, M. L., Teng, J. L. L. & Yuen, K. Y. Response to Evidence in favor of the essentiality of human cell membrane-bound ACE2 and against soluble ACE2 for SARS-CoV-2 infectivity. Cell 185, 1840-1841, doi:10.1016/j.cell.2022.05.005 (2022).
4 Kornilov, S. A. et al. Plasma levels of soluble ACE2are associated with sex, Metabolic Syndrome, and its biomarkers in a large cohort, pointing to a possible mechanism for increased severity in COVID-19. Crit Care 24, 452, doi:10.1186/s13054-020-03141-9 (2020).
5 Wallentin, L. et al. Angiotensin-converting enzyme 2 (ACE2) levels in relation to risk factors for COVID-19 in two large cohorts of patients with atrial fibrillation. Eur Heart J 41, 4037-4046, doi:10.1093/eurheartj/ehaa697 (2020).
6 Holman, N. et al. Risk factors for COVID-19-related mortality in people with type 1 and type 2 diabetes in England: a population-based cohort study. Lancet Diabetes Endocrinol 8, 823-833, doi:10.1016/S2213-8587(20)30271-0 (2020).
7 Bunyavanich, S., Do, A. & Vicencio, A. Nasal Gene Expression of Angiotensin-Converting Enzyme 2 in Children and Adults. JAMA 323, 2427-2429, doi:10.1001/jama.2020.8707 (2020).
8 Guo, X. et al. Regulation of age-associated insulin resistance by MT1-MMP-mediated cleavage of insulin receptor. Nat Commun 13, 3749, doi:10.1038/s41467-022-31563-2 (2022).
9 Chow, C. F. W. et al. Body weight regulation via MT1-MMP-mediated cleavage of GFRAL. Nat Metab 4, 203-212, doi:10.1038/s42255-022-00529-5 (2022).
10 Talmi-Frank, D. et al. Extracellular Matrix Proteolysis by MT1-MMP Contributes to Influenza-Related Tissue Damage and Mortality. Cell Host Microbe 20, 458-470, doi:10.1016/j.chom.2016.09.005 (2016).
11 Chan, K. M. et al. MT1-MMP inactivates ADAM9 to regulate FGFR2 signaling and calvarial osteogenesis. Dev Cell 22, 1176-1190, doi:10.1016/j.devcel.2012.04.014 (2012).
12 Wong, H. L. et al. MT1-MMP sheds LYVE-1 on lymphatic endothelial cells and suppresses VEGF-C production to inhibit lymphangiogenesis. Nat Commun 7, 10824, doi:10.1038/ncomms10824 (2016).
13 Jin, G. et al. MT1-MMP cleaves Dll1 to negatively regulate Notch signalling to maintain normal B-cell development. EMBO J 30, 2281-2293, doi:10.1038/emboj.2011.136 (2011).
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in